Hui Miao,
Cong-xiu Miao ![]() ,
Jing Han,
Na Li
,
Jing Han,
Na Li
For correspondence:- Cong-xiu Miao Email: miahui133494@163.com Tel:+863553128552
Received: 3 December 2015 Accepted: 1 August 2016 Published: 30 September 2016
Citation: Miao H, Miao C, Han J, Li N. Anti-hyperprolactinemia mechanism of Radix bupleuri extract in rats. Trop J Pharm Res 2016; 15(9):1903-1907 doi: 10.4314/tjpr.v15i9.13
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To determine the mechanism underlying the anti-hyperprolactinemia effects of Radix bupleuri extract (RBE) in rats.
Methods: Rats were divided into six groups (n=10 each group): healthy controls, untreated hyperprolactinemic rats, hyperprolactinemic rats treated with bromocriptine (0.6 mg/kg), and hyperprolactinemic rats treated with RBE (4.8, 9.6, or 19.2 g/kg). After 30 days, hypothalamic protein levels of dopamine D2 receptor, protein kinase A (PKA), and cyclic adenosine monophosphate (cAMP) were determined.
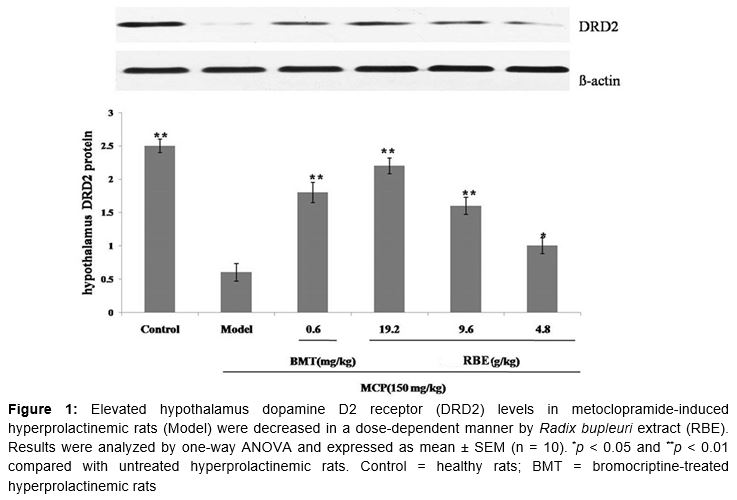
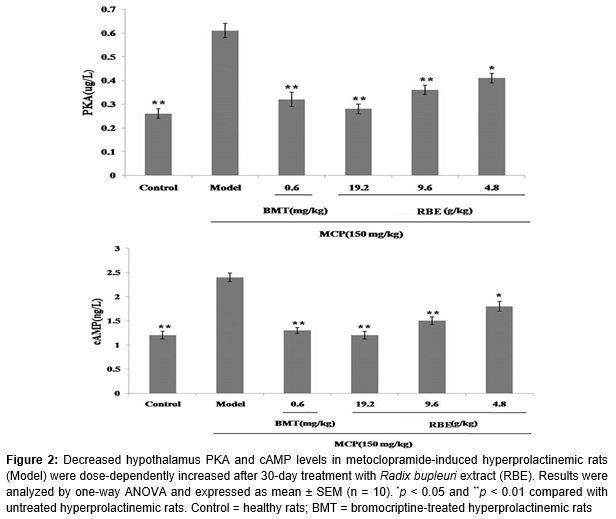
Results: Dopamine D2 receptor levels were lower in untreated hyperprolactinemic rats than in healthy controls (p < 0.01), but this decrease was attenuated by RBE (p < 0.05). Elevated PKA levels in untreated hyperprolactinemic rats (0.61 ± 0.04 µg/ml, p < 0.01) were decreased by RBE (4.8 g/kg, 0.42 ± 0.03 µg/ml, p < 0.05; 9.6 g/kg, 0.33 ± 0.02 µg/ml, p < 0.01; 19.2 g/kg, 0.27 ± 0.03 µg/ml, p < 0.01). Similarly, elevated cAMP levels in hyperprolactinemic rats (2.4 ± 0.4 ng/ml) were decreased by RBE (4.8 g/kg, 1.8 ± 0.3 ng/ml, p < 0.05; 9.6 g/kg, 1.5 ± 0.3 ng/ml, p < 0.01; 19.2 g/kg, 1.2 ± 0.2 ng/ml, p < 0.01).
Conclusions: RBE anti-hyperprolactinemia activity is mediated by dopamine D2 receptor signaling via the cAMP/PKA pathway.
Introduction
Hyperprolactinemia is a heterogeneous disorder characterized by elevated blood prolactin level. This disorder of the hypothalamic-pituitary axis can have a physiological cause (e.g., pregnancy), pathological cause (e.g., tumor), or unknown cause (i.e., idiopathic hyperprolactinemia). In different patients, similarly elevated prolactin levels can result in severe clinical manifestations or no symptoms at all [1]. Hyperprolactinemia is more common in women than in men. Its prevalence in an unselected adult population is 0.4 % but has been reported as 5 % in women visiting a family planning clinic, 9 % in women with adult-onset amenorrhea, and 17 % in women with polycystic ovary syndrome [2,3]. The two most commonly prescribed drugs for the treatment of hyperprolactinemia are bromocriptine and cabergoline. These medications are dopamine receptor agonists with similar characteristics and adverse effects (e.g., headache, nausea, and vomiting), but the frequency and severity of adverse effects appear to be lower for cabergoline. Although both drugs are effective in treating hyperprolactinemia, 12 % patients cannot tolerate bromocriptine [4].
Traditional Chinese medicine uses Radix bupleuri extract (RBE) to treat hyperprolactinemia without the toxic side effects associated with conventional anti-hyperprolactinemia agents. In a previous study, we found that RBE decreased prolactin levels in hyperprolactinemic mice [5]. However, the mechanism underlying the anti-hyperprolactinemia effects of RBE require further investigation.
Most conventional anti-hyperprolactinemia agents decrease prolactin secretion through the hypothalamic dopaminergic system, and these effects are mediated via the cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA) pathway [6]. In this study, we hypothesized that RBE alleviates hyperprolactinemia through modulation of the dopamine D2 receptor. We investigated the effects of RBE on protein levels of dopamine D2 receptor, PKA, and cAMP in the hypothalamus of hyperprolactinemic rats.
Methods
Preparation of RBE
Samples of Radix bupleuri were collected in May 2015 in Bozhou City, Anhui Province, China. Taxonomic identification of the plant was performed by Professor Pei Chen of Changzhi Medical College (Shanxi, China). A voucher specimen (No. RBE 201505034) was deposited in Changzhi Medical College for future reference.
One batch of R. bupleuri was dried in an oven. An aqueous extract of RBE was prepared by steeping the dried R. bupleuri in hot water (60 °C) three times (1 h each time). The extract was dried first in an oven and then freeze-dried. The yield was 62.5 % (1 g powder was obtained from approximately 1.6 g crude sample).
Animals
Female Wistar rats, weighing 200–220 g, were obtained from Shanxi Center for Disease Control and Prevention (Taiyuan, Shanxi). The animals had free access to food and water and were allowed to acclimatize for at least 1 week before experimental procedures. All experiments were approved by the Animal Care and Use Committee of Zhejiang Cancer Hospital (approval ref no. 20120622) and were carried out in compliance with Directive 2010/63/EU on the handling of animals used for scientific purposes [7].
The rats were treated with metoclopramide (150 mg/kg administered intraperitoneally daily for 10 days), a dopamine inhibitor widely used to generate animal models of hyperprolactinemia [8,9]. A total of 60 rats were divided into six groups (n = 10 per group): rats that did not receive metoclopramide (healthy controls), rats that received metoclopramide only (untreated hyperprolactinemic rats), hyperprolactinemic rats treated with 0.6 mg/kg bromocriptine (positive controls), and hyperprolactinemic rats treated with a high, medium, or low dose of RBE (19.2 g/kg, 9.6 g/kg, or 4.8 g/kg, respectively). The drugs were dissolved in 2 mL water and administered by intragastric gavage; healthy control and untreated hyperprolactinemic rats received 2 mL distilled water. RBE dosages were calculated from the human dosage based on body surface area. The mice were treated for 30 days.
Western blot analysis
To determine hypothalamic dopamine D2 receptor protein levels after the 30-day treatment period, rat hypothalamus tissues were homogenized in RIPA buffer with protease inhibitor (cOmplete EDTA-free protease inhibitor cocktail tablets; Roche Applied Science, Mannheim, Germany) and centrifuged at 3000 × g for 15 min at 4 °C. The supernatant was centrifuged at 12,000 × g for 20 min at 4 °C, and the samples were then diluted in RIPA buffer and loading buffer. After separation by 12 % sodium dodecyl sulfate polyacrylamide gel electrophoresis, the proteins were transferred onto polyvinylidene difluoride membranes (Merck). Nonspecific protein-binding sites were blocked by incubating with phosphate-buffered saline containing 0.1 % Tween-20 and 5 % fat-free milk for 1 h at room temperature, followed by incubation with primary antibodies against dopamine receptor D2 (1:1000) and ß-actin (1:5000), which served as a loading control. The membranes were then incubated with lumiGLO reagent (Cell Signaling Technology) and exposed to x-ray film (Kodak, Stamford, CT, USA). Protein bands were quantified by ImageJ (National Institutes of Health, Bethesda, MD, USA). Relative D2 receptor protein levels were determined by normalization to ß-actin.
Enzyme-linked immunosorbent assay
To determine hypothalamic cAMP and PKA protein levels after the 30-day treatment, rat brains were stripped of membranes and blood vessels, homogenized, and then centrifuged for 15 min at 3000 r·min-1. Protein levels of PKA and cAMP in the supernatant were then determined by enzyme-linked immunosorbent assay (ELISA; Nanjing Jiancheng Biological Technology Co., Ltd, China).
Statistical analysis
Treatment groups were compared by one-way analysis of variance (ANOVA), followed by Student–Newman–Keuls post hoc test for multiple comparisons. Data are expressed as mean ± SEM; p < 0.05 was considered statistically significant.
Results
Effects of RBE on hypothalamus dopamine D2 receptor levels in hyperprolactinemic rats
Results of western blot analysis showed that hypothalamus dopamine D2 receptor protein levels were considerably lower in untreated hyperprolactinemic rats than in healthy controls (p < 0.01). As expected, this decrease was attenuated by 30-day treatment with 0.6 mg/kg bromocriptine (positive control, p < 0.01). Treatment with RBE resulted in a dose-dependent decrease in dopamine D2 receptor levels (4.8 g/kg, p < 0.05; 9.6 g/kg, p < 0.01; 19.2 g/kg, p < 0.01) ().
Effect of RBE on hypothalamus PKA and cAMP levels in hyperprolactinemic rats
Results of ELISA showed that hypothalamus levels of PKA and cAMP were significantly higher in hyperprolactinemic rats compared with healthy controls (p < 0.01). This upregulation of PKA and cAMP protein expression was attenuated by treatment with bromocriptine (p < 0.01). Treatment with RBE resulted in a dose-dependent decrease in PKA and cAMP levels (4.8 g/kg, p < 0.05; 9.6 g/kg, p < 0.01; 19.2 g/kg, p < 0.01) ().
Discussion
In a previous study, we found that RBE significantly decreased prolactin levels in a mouse model of hyperprolactinemia. In the present study, we investigated the mechanism underlying the anti-hyperprolactinemia effects of RBE and found that RBE significantly increased dopamine D2 receptor protein levels and significantly decreased cAMP and PKA protein levels in the hypothalamus of hyperprolactinemic rats.
Dopamine receptors are members of the G-protein coupled receptor family [10-12] and are classified on the basis of their effect on adenylyl cyclase, which regulates the cAMP-PKA pathway.
D1-like dopamine receptors (D1 and D5) couple primarily to Gαs and increase intracellular levels of the second messenger cAMP and upregulate the activity of PKA. By contrast, D2-like dopamine receptors (D2, D3, and D4) couple to Gαi/o and decrease levels of cAMP, thereby suppressing PKA activity [13–15].
In the adenohypophysis, which secretes prolactin, the predominant dopamine receptor is the D2 receptor [16-18]. In a pituitary cell line transfected with the gene encoding the dopamine D2 receptor, treatment with dopamine decreased intracellular cAMP levels and inhibited prolactin secretion [19,20]. Similarly, most conventional anti-hyperprolactinemia agents are D2 receptor agonists, which decrease prolactin secretion through cAMP/PKA signaling. In this study, RBE increased dopamine D2 receptor levels and decreased cAMP and PKA levels in the hypothalamus of hyperprolactinemic rats significantly. These results suggest that the anti-hyperprolactinemic activity of RBE is mediated by dopamine D2 receptor and cAMP/PKA signaling.
Conclusion
Radix bupleuri extract decreases prolactin secretion in hyperprolactinemic rats via dopamine D2 receptor and cAMP/PKA signaling. This mechanism is similar to that of conventional treatments for hyperprolactinemia, without the toxic side effects. This traditional Chinese medicine may, therefore, be useful for the treatment of patients with hyperprolactinemia.
Declarations
Acknowledgement
References
Archives


News Updates