Araya Supawat,
Supawadee Srisuwan,
Jintana Sattayasai ![]()
For correspondence:- Jintana Sattayasai Email: sjinta@kku.ac.th Tel:+6643348397
Received: 18 February 2016 Accepted: 12 June 2016 Published: 31 July 2016
Citation: Supawat A, Srisuwan S, Sattayasai J. Oral glutamate intake reduces acute and chronic effects of ethanol in rodents. Trop J Pharm Res 2016; 15(7):1493-1498 doi: 10.4314/tjpr.v15i7.20
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To assess the effects of oral glutamate intake on acute motor effects and chronic intake of ethanol in rodents.
Methods: The acute effects of ethanol on motor function were studied in ICR mice by giving 2 or 6 g/kg of ethanol 2 h after distilled water or 2.5 g/kg glutamate per os. Thirty minutes after ethanol treatment, behavioral assays, including rotarod tests and foot print analysis were monitored. In chronic ethanol treatment, male Wistar rats were trained to consume ethanol-sucrose solution during a 2-h period daily, starting with 2 % ethanol/10 % sucrose and gradually increasing to 10 % ethanol/5 % sucrose solution over 56 days. After training session, the drug treatment phase was done for 10 days. The animals were force-fed 50 mg/kg/day topiramate or 2.5 g/kg/day glutamate 2 h before ethanol treatment sessions. Each day, ethanol intake, water intake, food intake and body weight were recorded.
Results: Mice that received 2 or 6 g/kg of ethanol orally, showed a significant reduction in time on the rod in the rotarod test and a significant increase in both forelimb and hindlimb stride lengths when compared to control. Oral treatment with 2.5 g/kg of glutamate reversed the acute motor effects of ethanol. In chronic ethanol treatment, the intake of 10 % ethanol/5 % sucrose, accessible for 2 h, was significantly decreased in rats treated with either topiramate or glutamate.
Conclusion: These results provide evidence that oral glutamate administration help to reduce the acute motor effects of ethanol in mice and ethanol intake in the chronic ethanol drinking rats.
Introduction
Glutamate, the principal excitatory amino acid neurotransmitter present in the mammalian brain, is one of the neurotransmitters responsible for neuronal effects of ethanol in both acute and chronic alcohol consumption. In acute alcohol consumption, all glutamate receptors, including N-methyl-D-aspartate (NMDA) and L-alpha-amino-3-hydroxy-5-methyl-isoxazole-4-propionic acid ionotropic (AMPA) receptors are inhibited [1]. Together with increased inhibitory neurotransmission through gamma-aminobutyric acid (GABA), acute alcohol exposure leads to a sedated state and impaired motor coordination in drinkers [1,2]. On the other hand, chronic use of alcohol seems to upregulate NMDA receptor expression in the brain and leads to alcohol dependence and relapse-like alcohol drinking [1,2]. Glutamate also plays an important role in the reward system of alcohol addiction [3,4,5]. Increased extracellular glutamate levels were observed in the reward system during ethanol withdrawal [6]. Currently, several studies have focused on the investigation of glutamatergic agents in maintaining glutamate homeostasis, modifying alcohol’s effects, and reducing alcohol intake [7,8]. Acamprosate, one of the approved treatments for alcoholism, is proposed to exert at least part of its effsct by altering glutamatergic function [9,10]. Topiramate, an anticonvulsant, that inhibits glutamate release and blocks AMPA/kainate receptors, can also be used in the treatment of alcohol dependence [11].
Glutamate is found in abundant amounts in many natural and prepared foods as a food flavoring agent for the umami taste. Recently, it was shown that monosodium glutamate at 2.5 or 4 g/kg per os could markedly elevate the glutamate level in the brain and other organs in mice and reached peak values at 90 - 120 min [12]. Thus, when drinking alcohol with food as a source of glutamate, interactions between ethanol and glutamate might occur. This study was aimed to investigate the effects of oral administration of glutamate on the acute effects of ethanol on motor functions and alcohol intake in alcohol addiction.
Methods
Experimental animals
In this study, male outbred Mlac:ICR mice, 6-8 wks old, weighing 25–35 g, and male outbred Wistar rats, 6-8 wks old, weighing 250–350 g, were purchased from the National Laboratory Animal Center, Mahidol University, Thailand. The animals had free access to a commercial diet (CP, Thailand) and water, and were housed in a room with a 12-h light: 12-h dark cycle under controlled temperature and humidity. All animal experiments were strictly in accordance with international ethical guidelines [13] and Ethics of Animal Experimentation of National Research Council of Thailand concerning the Care and Use of Laboratory Animals. The experimental protocols also received the approval of Animal Ethics Committee of Khon Kaen University (approval no. 0514.1.12.2/78).
Chemicals and reagents
Ethanol (BDH Prolabo Chemicals, EC), sucrose (Ajax Fine Chem Pty Ltd, New Zealand), topiramate (Topamax®, Janssen Pharmaceuticals, Inc. Toronto, Canada) and L-glutamic acid (Sigma-Aldrich, USA) were used in this study. The chemicals were dissolved in distilled water on the day of the experiment and given to the animals in a dose of 0.05 ml/10 g body weight. Doses of ethanol used were expressed as g/kg.
Acute ethanol treatment
Each of the mice were force fed two times 2 h apart. First, distilled water or 2.5 g/kg glutamate [12] was administered followed by distilled water or ethanol (2 or 6 g/kg) [14]. Thirty minutes after the second treatment, the rotarod tests and foot print analyses were performed.
Rotarod test
The rotarod test was done to assess motor coordination and balance in mice. The AccuRotor Rotarod (Accuscan Instruments, Columbus, OH) was used for fixed-speed rota-rod (FSRR) tests. Mice were placed with the forepaws on bars (diameter 2.5 cm) which were turning at 12 rpm and the time that the animal remained on the rod was recorded [14]. The maximum observed time was set at 60 s.
Foot print analysis
Foot print analysis was used to assess ataxia and gait abnormalities in mice as described earlier [15]. Animals were made to walk along a 60 cm long, 7 cm wide runner with 10 cm walls by lighting a lamp at the start and placing a dark box at the end. The runner was lined with white paper, and the fore and hind paws of the animals were dipped in red and blue nontoxic colors to record the footprints. Six middle steps of a series of steps were analyzed and the distances between two forelimb and two hindlimb pawprints were measured as forelimb stride length (FSL) and hindlimb stride length (HSL), respectively.
Assessment of chronic ethanol consumption in rats
In this chronic study, male Wistar rats were treated as described earlier [16] and topiramate, an antiepileptic agent that reduces ethanol consumption [11] was used as a positive control. Animals were individually housed and received ad libitum access to standard rodent chow and water. The experiments were performed during the light cycle. In the training phase, rats were given free access to ethanol in sucrose solutions during two-hour sessions at 10:00-12:00 am of each day for 56 days. Ethanol–sucrose solutions were presented in 25 mL graduated plastic tubes with rubber stoppers and metal sipper tubes were inserted through the cage. The concentration of ethanol in these solutions was systematically increased, while the concentration of sucrose was decreased over a 56-day period. The ethanol-sucrose solution given to the animals were 2 % ethanol/10 % sucrose solution on days 1-11, 5 % ethanol–5 % sucrose solution on days 12 - 24, 8 % ethanol/5 % sucrose solution on days 25 - 38 and 10 % ethanol/5 % sucrose solution on days 39 - 56 which continued for another 10 days for the treatment phase. The 10th day drug treatment phase began after animals had completed 8 weeks (56 days) of training on drinking. Animals were randomly assigned to control (distilled water), topiramate at 50 mg/kg/day and glutamate at 2.5 g/kg/day treatment groups with 6 animals in each group. On each day of the drug treatment phase, 2 h before the ethanol session, animals received intragastrically distilled water, 50 mg/kg topiramate or 2.5 g/kg glutamate, once daily. After that, a 2-h session of free access to a 10 % ethanol–5 % sucrose solution was initiated. On each day, the amount of ethanol intake (g/kg) in the two-hour session, water intake (ml/day) and food intake (g/day) were recorded. Animal body weights were recorded daily during the whole experiment to monitor drug-induced body weight changes.
Statistical analysis
The data are presented as means ± standard error of means (SEM). For the acute effects of ethanol on motor functions, one-way analysis of variance (ANOVA) followed by Tukey post-hoc test) and Student t-tests were used. Results were considered significant at p < 0.05. In the assessment of chronic ethanol consumption in rats, two-way ANOVA with groups and times as factors, were performed.
Results
Effect of glutamate on acute ethanol treatment
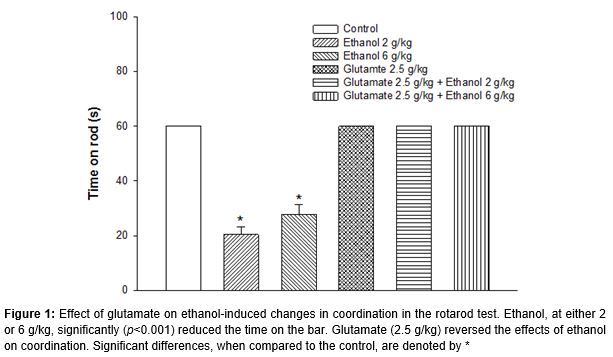
In the rotarod test, the control animals on water-only showed normal coordination and could stay on the rod for over 60 sec. Ethanol, at either 2 or 6 g/kg, significantly (p < 0.001) decreased the time on rod when compared to the control (). Mice that received 2.5 g/kg glutamate had normal coordination and could stay on rod for over 60 sec. Time on rods of mice that received glutamate with either 2 or 6 g/kg of ethanol were comparable to the control which suggested that glutamate could completely reverse the effects of ethanol.
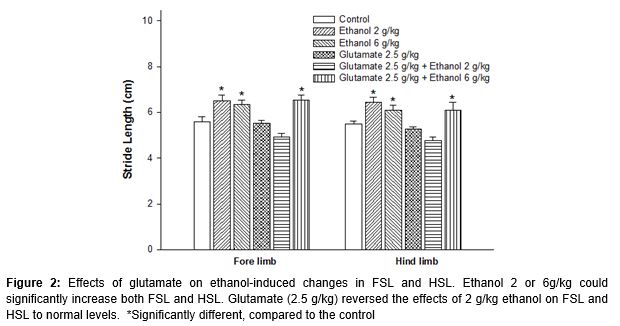
Foot print analysis showed that the control animals used only the front parts of the paws to walk on and a narrow-based stance with close proximity forelimb and hindlimb footprints with 5.71 ± 0.48 and 5.30 ± 0.49 cm of forelimb and hindlimb stride lengths. The stepping patterns of animals that received either doses of ethanol were different from the control animals. Animals treated with ethanol, at either 2 or 6 g/kg, walked on their whole paw with a significant increase in FSL (p = 0.037, 0.043) and HSL (p = 0.010, 0.050) when compared to the control (). Mice that received 2.5 g/kg glutamate could reverse the effects of 2 g/kg, but not 6 g/kg, of ethanol on FSL and HSL to the control length.
Effect of glutamate on chronic ethanol consumption
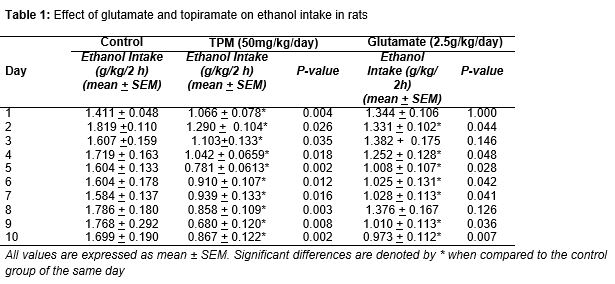
At the end of the training phase, the mean daily intake of ethanol in g/kg for each treatment group for the baseline phase, was of 1.529 g/kg/2-h session (± 0.088 SEM). During the 10 days of the treatment phase, the control animals showed no change in the amount of ethanol intake. A significant reduction of ethanol intake that was seen in topiramate-treated group, when compared to the control, can be seen from day 1 to day 10 (). Glutamate treatment, at 2.5 mg/kg/day, also significantly reduced the amount of ethanol intake from day 2 to day 10 and the effects were comparable to topiramate at 50 mg/kg/day. A two-way repeated measures ANOVA showed a significant effect of treatment [F(3, 239) = 16.618, p < 0.001], without any significance in treatment x day interactions (p = 0.869).
Significant reduction of ethanol intake (g/kg/two-hour session) can be seen in rats treated with topiramate (50 mg/kg/day) or glutamate (2.5 g/kg/day) from day 1 or day 2 of the treatment phase and was still prominent at the end of the experiment.
No significant differences of the mean volume of water, food intake or change of body weight could be seen in all treatment groups when compared to the control (data not shown).
Discussion
Recently, it has been shown that monosodium glutamate (MSG), the primary component of umami taste, can be used as a substitute to initiate ethanol drinking in both C57BL/6J and DBA/2J mice [17]. Interestingly, this study is the first to show that intragastric administration of glutamate could reduce both the acute effects of ethanol on motor functions and chronic ethanol intake.
Generally, ethanol modulates synaptic efficacy in many brain areas, including the cerebellum and cortical regions [18,19] and causes motor impairment from the inhibition of ionotropic glutamate receptors and enhancing the inhibitory action of GABA [20,21]. It is noted that glutamate could be detected in the brain with peak concentrations at approximately 2 h after oral administration of 2.5 g/kg glutamate [17]. In the present study, oral administration of glutamate, 2 h before ethanol, might have increased the glutamate levels in the brain, activated glutamate receptors and helped reduce the acute effects of ethanol on motor functions.
In the chronic ethanol consumption model, topiramate treatment clearly reduced the ethanol intake of the rats. It has been shown previously that topiramate, by facilitation of inhibitory GABA effects and antagonism of AMPA/kainate glutamate receptors can decrease ethanol reinforcement and the propensity to drink [22,23]. Recently, glutamate was able to induce more ethanol drinking in C57BL6/J and DBA/2J mice and the authors suggested that the umami effect of glutamate is responsible for the effect [17]. In the present experiment, glutamate administered by an intragastric tube, bypassed the umami taste, reduced ethanol intake in rats with a history of chronic alcohol consumption. It is interesting to note that changes in the balance between synaptic and extrasynaptic glutamate levels in the brain in turn influence signaling through pre- and post-synaptic glutamate receptors, and thus affect synaptic plasticity and circuit-level activity [24].
In chronic exposure to cocaine, a decrease in extrasynaptic glutamate concentrations in the nucleus accumben (NAc) and a decreased tone of presynaptic inhibitory mGluR2/3 glutamate receptors was observed [24]. MGluR2/3 glutamate receptors are inhibitory to glutamatergic neurotransmission; the decreased tone of these receptors leads in turn to enhanced glutamate release in the prefrontal cortex-NAc projection and activates drug-seeking behavior [25]. Giving glutamate orally might increase extrasynaptic glutamate level in the brain [17,26] and reduce ethanol-seeking behavior.
The results from this study suggest that, glutamate, as one of the common ingredients found in all kinds of foods, might exert a very interesting interaction with ethanol in either its acute or chronic effects.
Conclusion
These results provide evidence that oral glutamate administration reduces acute motor effects of ethanol and ethanol intake in the chronic ethanol drinking rat model.
Declarations
Acknowledgement
References
Archives


News Updates