Peng Ding1,2,
Xi-Miao Xu3,
Liang-Zhu Teng1,
Ji-Jun Xu2,
Xu-Yuan Zhou2 ![]()
For correspondence:- Xu-Yuan Zhou Email: zhaoxuyuanews@hotmail.com Tel:+866325511021
Received: 4 September 2015 Accepted: 16 December 2015 Published: 29 January 2016
Citation: Ding P, Xu X, Teng L, Xu J, Zhou X. Tricyclic neovibsanin scaffold inhibits glioma by rargeting glioma-initiating cells. Trop J Pharm Res 2016; 15(1):33-38 doi: 10.4314/tjpr.v15i1.5
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the effect of tricyclic neovibsanin scaffold (TCNS) on cell viability, colony formation capacity and induction of apoptosis in glioma cells.
Methods: 3-(4, 5-Dimethylthiazol-2-yl) 2, 5-diphe¬nyltetrazolium bromide (MTT) assay was used to analyze the effect of TCNS on cell proliferation. Light microscopic examination of giemsa solution stained cells was used to calculate the number of colonies with > 50 cells. Flow cytometry using a flow cytometer, while apoptosis detection kit were used to analyze induction of apoptosis.
Results: T CNS treatment significantly inhibited the viability of U138 NS and U138 AC cells in a concentration-dependent manner (p < 0.05). TCNS caused 86 % reduction in the capacity of U138 NS cells to form colonies and led to significant induction of apoptosis. The activation of caspase 3 and ex
Conclusion: Thus, TCNS treatment significantly inhibits the viability of glioma cells and colony formation, but induces apoptosis and increases the median survival of mice. Hence, TCNS may be of therapeutic value for the treatment of glioma.
Introduction
Glioma is the most frequently detected primary brain tumor in adults and has a median survival time of around 3 months for patients availing no treatment [1]. The currently used treatment strategies for glioma, include chemotherapy, radiotherapy and surgical resection. However, because of the enhanced rate of proliferation and aggressive invasive and metastasis potential the rate of glioma prognosis is very poor. Glioma initiating cells (GICs) comprise a group of cells which are associated with beginning and progress of brain tumor [2 3]. These cells are bestowed with the capacity of self-renewal and multiplication. GICs are resistant to various anticancer agents, leading to the failure of cancer treatment strategies [4,5]. Therefore the development of novel and efficient treatment strategies for glioma inhibition are required [6-10].
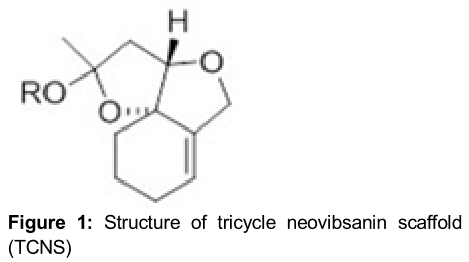
Neovibsanins are diterpenoid compounds present in plants and possess polyfunctional structures. They were isolated by Fukuyama’s research group in 1996 during photochemical investigation of the extract of the shrub, Viburnum awabuki [11]. Later, these compounds were found to promote neurite outgrowth activity in PC12 cells. Therefore, it is believed that neovibsanin-type compounds can be of vital importance for the treatment of neurological disorders, including Alzheimer’s disease [12]. It was observed that not only neovibsanins but also their synthetic derivatives exhibit neurite outgrowth activities. There is hardly any effect of stereochemistry on the biological activity of neovibsanins [13]. The tricyclic neovibsanin scaffold showed neurite outgrowth activity similar to that of the parent neovibsanin [14]. The present study was performed to investigate the effect of tricyclic neovibsanin scaffold () on cell proliferation, colony formation capacity and induction of apoptosis in glioma cells. It was observed that TCNS inhibits cell proliferation and colony formation and induces apoptosis in glioma cells.
Methods
Animals
A total of 10 BALB/c mice, 8 - 10 week old and 22 ± 2 g in weight were purchased from the Experimental Animal Center of Shandong University (Jinan, China). The animals were acclimatized to the laboratory conditions one week before the start of experiment and were maintained at 24 oC with free access to water and standard laboratory chow. All the experimental procedures were performed according to the guidelines of Laboratory Animal Care Committee of Sun Yat-sen University (Guangzhou, China). Approval was taken from the ethics and review committee for the animal studies at Laboratory Animal Care Committee of Sun Yat-sen University (approval ref no. 179 - 2008).
Reagents
Tricyclic neovibsanin scaffold (TCNS) was synthesized using the reported protocol [14] and dissolved in DMSO to prepare the stock solution. Fetal bovine serum (FBS) and Dulbecco's modified Eagle's medium (DMEM) were purchased from Life Technologies (Gaithersburg, MD, USA). Dimethyl sulphoxide and other common chemicals were obtained from Sigma (St. Louis, MO, USA).
Cell culture
Glioma cell line, GL261 was purchased from Shanghai Institute of Biochemistry and Cellular Biology Chinese Academy of Sciences (Shanghai, China). The cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10 % fetal bovine serum and antibiotics. Then the cells were maintained in a humidified atmosphere at 37 oC with 5 % CO2 and 95 % air.
Immunocytochemistry analysis
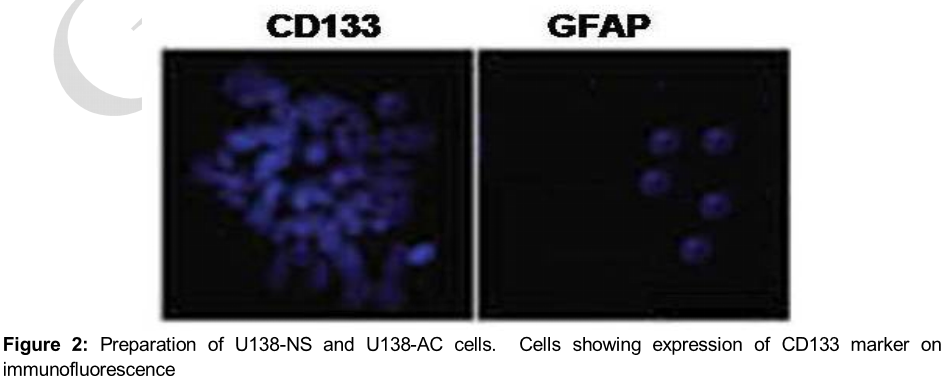
On to the poly L lysine coated coverslips cells were dispersed and the slides were fixed with paraformaldehyde. After 45 min normal goat serum was applied to block the cell samples followed by overnight incubation with anti-mouse CD133/1 monoclonal and GFAP polyclonal primary antibodies. The cells were subjected to Cy3 conjugated goat anti rabbit secondary antibody staining after washing with PBS. Following 45 min incubation the cells were counterstained using 100 mg/mL DAPI for 20 min. A confocal laser scanning microscope (Leica, Mannheim, Germany) was used for analysis of the cell nuclei.
Assessment of cell viability
For determination of cell viability colorimetric 3-(4, 5-dimethylthiazol-2-yl) 2, 5-diphe-nyltetrazolium bromide (MTT) assay was used. Briefly, the cells at a density of 2 x 105 cells per well were dispersed onto 96 well culture plates and treated with various concentrations of TCNS for 24 h. After incubation, the cells were rinsed two times in phosphate buffered saline (PBS) followed by incubation for 4 h with MTT (5 mg/mL). Dimethyl sulphoxide (150 µl) was added to each well for dissolving the insoluble blue formazan crystals formed. Absorbance was measured for each well at 565 nm with an enzyme linked immunosorbent assay plate reader (DR-200Bs; Bio Rad Laboratories, Hercules, CA, USA).
Colony formation assay
In order to determine the effect of TCNS on the capacity to form colonies, the cells were distributed at a density of 2 x 105 cells per ml on to 12-well plates. The cells were exposed to TCNS, taxol or DMSO and incubated for 5 days. After incubation, the cells were subjected to crystal violet solution staining and the number of colonies formed was counted using an Olympus CX22 microscope (Olympus Corp, Inc, Tokyo, Japan).
Flow cytometric analysis of apoptosis
The cells after treatment with TCNS were harvested using trypsinization and rinsed in phosphate buffered saline twice. The cells were then re-suspended in binding buffer at a concentration of 2.5 x 106 cells per mL, and 5 μL each of annexin-V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) were added to the cell suspension and incubation was continued for 15 min under dark atmosphere. FACScalibur Flow Cytometer (BD Biosciences, San Jose, CA, USA) was used to measure the apoptotic cell percentage.
Western blot analysis
TCNS treated cells were harvested and washed twice with PBS. The cells were the lysed for 40 min in lysis buffer (50 mM Tris, pH 7.4, 1 % Triton X 100, 150 mM NaCl, 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride, 1 mM ethylene glycol tetraacetic acid, 10 μg/mL aprotinin, 1 mM sodium orthovandate, 10 μg/mL leupeptin and 1 mM NaF). Cell lysates were centrifuged at 12,000 x g for 45 min at 4 oC. Electrophoresis using 10 % sodium dodecyl sulfate polyacrylamide gel was used to resolve the proteins from each sample. The proteins were transferred onto a nitro-cellulose membrane and incubated with primary antibodies using blocking buffer. After incubation with primary antibodies the membrane was washed again with PBS. The membrane was then incubated with horseradish peroxidase conjugated goat anti-rabbit IgG (Wuhan Boster Biological Technology, Ltd) secondary antibodies. β-actin was examined on the same membrane and used as a loading control. The electrochemiluminesence reagent (GE Healthcare, Amersham, UK) was used to detect the antibody bound complexes. Optical density (OD) was determined for the relative levels of the target protein.
Xenograft tumor formation assays
TCNS treated and control cells were harvested and suspended in 200 μL of sterile saline at a density of 2 x 106 cells. Among two groups of mice with 5 mice in each group, one group was administered TCNS treated cells and the second group untreated cells subcutaneously. After 2 months of administration of cells, the survival time of the mice was compared.
Statistical analysis
All the data are expressed as mean ± SD (n = 3). One way analysis of variance and SPSS 16.0 software (SPSS, Inc., Chicago, IL, USA) were used for the analysis of differences in data. Differences were considered significant statistically at p < 0.05.
Results
U138 NS and U138 AC cells
U138 cells were induced to form neurospheres by culturing in a medium devoid of serum and containing basic fibroblast growth factor, epidermal growth factor, L glutamine and insulin. Formation of neurospheres was evident by the presence of CD133 cell marker (). Differentiation was induced by replacing the media every 4 days with DMEM supplemented with 10 % FBS. It was observed that the cells got adhered and started to differentiate forming U138 AC cells.
Inhibition of U138 NS and U138 AC cell viability by TCNS
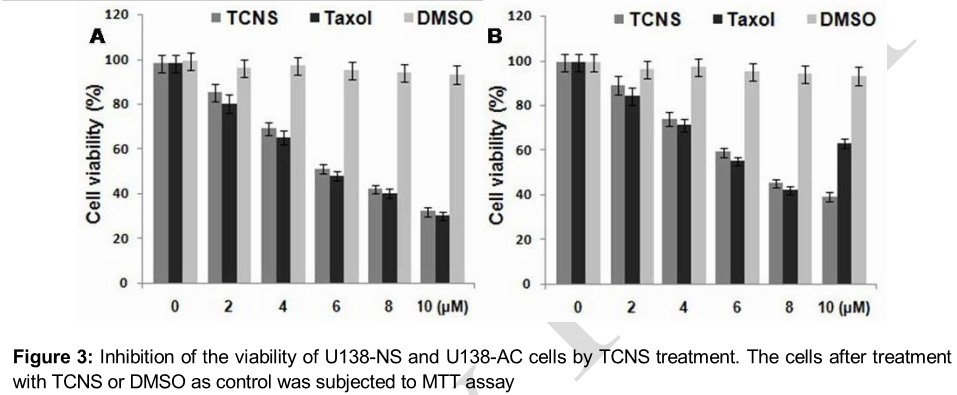
The results of MTT assay revealed that TCNS treatment inhibited the viability of U138 NS and U138 AC cells in concentration dependent manner. Among the range of TCNS concentrations from 1 to 10 µM used the minimum inhibitory concentration of TCNS against U138 NS and U138 AC cells was 4 and 6 µM, respectively ().
Inhibition of colony formation in U138 NS cells by TCNS
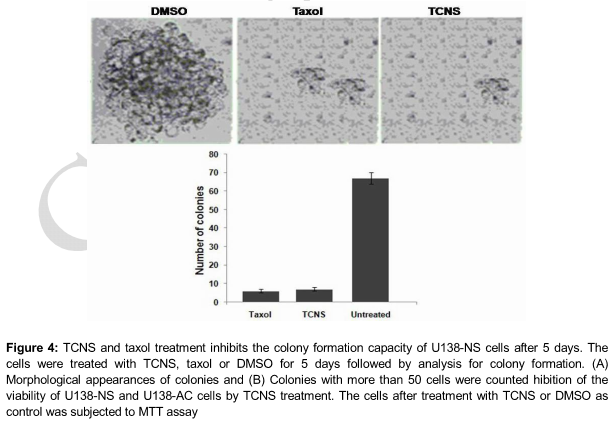
Exposure of U138 NS cells to 5 µM concentration of TCNS for 5 days caused a significant decrease in the capacity to form colonies. Comparison of the inhibition in colony formation with taxol, a known anticancer agent revealed that TCNS at 5 µM concentration exhibited similar inhibitory effect as that of taxol at 20 µM. The reduction in colony formation was 86, 84 and 5 % by TCNS, taxol and DMSO, respectively, after 5 days ().
Induction of apoptosis in U138 NS cells by TCNS
Exposure of U138 NS cells to TCNS at 5 µM concentration caused a significant increase in apoptotic cell population compared to the control cells (p < 0.05; A). The results from western blot analysis revealed that TCNS enhanced the activation of caspase 3 protein in U138 NS cells after 24 h. TCNS also induced the expression of Bax protein significantly following 24 h treatment (p < 0.05; B). The increase in the activation of caspase 3 and expression of Bax in TCNS and taxol treated cells was similar.
TCNS prolongs the median survival time of tumor bearing mice
Examination of the median survival time of rats revealed that TCNS caused a significant increase in the median survival time of mice compared to the untreated rats. The median survival times were 34, 33 and 22 days respectively for mice in normal, TCNS treated and untreated mice, respectively.
Discussion
Glioma initiating cells (GICs) comprise a group of cells possessing the ability of self-renewal and recapitulation. Natural products play an important role in the development of chemopreventive and chemotherapeutic agents used for treatment of various cancers [15]. Neovibsanins are diterpenoid compounds present in plants and possess polyfunctional structures. They were isolated by Fukuyama’s research group in 1996 during photochemical investigation of the extract of the shrub, Viburnum awabuki [11]. The present study was performed to investigate the effect of tricyclic neovibsanin scaffold on proliferation, colony formation, induction of apoptosis and increase in median survival of mice.
Cancer cells possess the ability to proliferate at an infinite rate, therefore, the inhibition of cell proliferation is an important treatment strategy for cancer therapy.
In the present study TCNS significantly inhibited the proliferation of glioma cells. Apoptosis is the programmed death of cells regulated by genes and imbalance between apoptosis and cell proliferation leads to various disorders, including carcinoma. Cancer cells are also resistant to apoptosis, and therefore, the induction of apoptosis in carcinoma cells by various agents can be of therapeutic importance for cancer treatment [16,17]. In the present study treatment of glioma cells with TCNS caused a significant increase in the induction of cell apoptosis.
For understanding the mechanism of TCNS induced apoptosis the expression of Bax and activation of cleaved caspase 3 were analyzed. Caspase 3 plays a vital role in the induction of cell apoptosis [18,19]. Activated Bax enters into the mitochondrial membrane leading to enhanced membrane permeability and release of cytochrome C [20]. Cytochrome C then binds with apoptotic protease activating factor 1 and ATP and ultimately results in an apoptotic cell [18]. In the present study, the levels of cleaved caspase 3 and Bax were significantly increased. Therefore, it was deduced that TCNS exerts its proapoptotic effects by increasing the levels of proapoptotic proteins. In addition, TCNS increases the median survival of mice significantly compared to the untreated mice.
Conclusion
TCNS significantly inhibits U138NS and U138AC cell viability, cell colony forming capacity, induces apoptotic cell death and increases median survival time of mice bearing glioma. Thus, TCNS may be of therapeutic importance in the treatment of malignant glioma.
References
Archives


News Updates