Hyun-Joo Chang1,
Hyun Kang2 ![]()
For correspondence:- Hyun Kang Email: hkang@dankook.ac.kr Tel:+82415501452
Received: 15 September 2015 Accepted: 28 December 2015 Published: 29 January 2016
Citation: Chang H, Kang H. Ulmus davidiana var japonica attenuates neuroinflammatory responses in activated bv2 murine microglial cells via nuclear factor-kappab signaling pathway. Trop J Pharm Res 2016; 15(1):19-26 doi: 10.4314/tjpr.v15i1.3
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the antioxidant and anti-inflammatory effects of Ulmus davidiana extract (UDE) in lipopolysaccharide (LPS)-stimulated BV-2 cells.
Methods: Antioxidant activity was measured using 1, 1-diphenyl-2-picryl-hydrazyl (DPPH) radical scavenging assay. Cell viability was evaluated using 3-(4, 5-dimethylthiazol-2-yl)-2, 5-
diphenyltetrazolium bromide (MTT) assay. BV-2 cells were stimulated with LPS to study protein ex
Results: UDE significantly inhibited DPPH-generated free radicals showing maximum inhibition at 40 μg/mL (p < 0.001). UDE alone did not exhibit any signs of cytotoxicity towards BV-2 cells up to 100 μg/mL concentration. The LPS-induced increase in the production of nitric oxide was concentration-dependently suppressed with half-maximal concentration (IC50) of 67.4 µg/ mL of UDE (p < 0.05 at 10 μg/mL, p < 0.01 at 20 μg/mL and p < 0.001 at 40 μg/mL, respectively). UDE also inhibited dose-dependently the LPS-induced increase in inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) ex
Conclusion: This study shows, for the first time, that UDE possesses antioxidant and anti-inflammatory effects and can be developed as a potential therapeutic agent for ameliorating macrophage-mediated inflammation.
Introduction
Ulmus davidiana var. japonica (UDE), from the family of Ulmaceae, is widely distributed in Korea, Japan, and China. The stem and root barks of this plant are traditionally used to treat diarrhea, edema, jaundice, hepatitis, and gastric cancer [1]. Its proven medicinal potency merits an elucidation of the mechanisms by which it works and this study will focus on the molecular activities of UDE on BV2 murine microglial cells.
UDE can be used to promote diuresis and to treat dampness [1] It is composed of various sesquiterpenes, triterpenes, lignans, and flavonoids [2]. Previous studies have shown that UDE or its isolated components have various effects, including anti-oxidation [2,3], anti-inflammatory [3], and possibly anti-angiogenic effect [4].
Neuro-inflammation mediated by microglial activation appears to play an essential role in the pathogenesis of various neuro-degenerative diseases such as Alzheimer’s disease, Parkinson’s disease and multiple sclerosis [5]. Microglia under activated conditions releases several inflammatory cytokines and toxic free radicals, which may contribute to the neurodegenerative process. Previous reports demonstrate that suppressive agents of the inflammatory mediator concentrations in activated microglia can attenuate the severity of neuro-degenerative diseases [6].
It is well documented that the pathophysiological condition in inflammatory disorders is initiated by a complex processes triggered by microbial endotoxins like LPS, which can directly activate microglia, thus triggering the production of inflammatory mediators, such as NO, COX-2, pro-inflammatory cytokines and leukotrienes [7]. Therefore, LPS-induced inflammatory mediators in vitro can be regarded as one of the essential tools for evaluating the deleterious consequences of excessive activation of microglia. Recent studies have shown that antioxidant and anti-inflammatory agents may suppress microglial activation and thus protect neuronal cell death [8].
In this study, the in vitro antioxidant potential and anti-neuroinflammatory properties of UDE were evaluated in LPS- and interferon-γ-stimulated BV2 murine microglial cells.
Methods
Preparation of U. davidiana extract
Dried plant material of U. davidiana purchased from a market specializing in herbs (Traditional herb market; Seoul, Korea). It was authenticated by Prof Chang-Kyu Oh, a taxonomist at Dankook University, South Korea authenticated. A voucher specimen (UD-DKU2015) was stored in Department of Medical Laboratory Science, Dankook University for future reference. To obtain the U. davidiana extract, the dried plant material was ground in a blender. The powdered material was extracted with 10 volumes (v/w) of 70 % ethanol at room temperature for 72 h and filtered. The residue was extracted with absolute ethanol at 1:10 ratio (w/v) for 2 h in a heating mantle at 70 - 80 °C, and the supernatant was filtered and concentrated in a vacuum evaporator system at 50 °C. For further fractionation, the extract (1000 mg) was partitioned into hexane, chloroform and ethyl acetate (EA) fractions to yield 267, 73 and 307 mg, respectively. The active EA fraction of U. davidiana (UDE) was re-dissolved in distilled water.
DPPH radical scavenging activity
This assay was based on the scavenging of stable 2, 2-diphenyl-1-picrylhydrazyl (DPPH, Sigma-Aldrich, St. Louis, MO, USA) radicals by the radical scavenging antioxidants in UDE. The radical scavenging capacity was evaluated by employing a reaction mixture constituted by aliquots of the UDE and a DPPH methanolic solution as described previously [8]. A sample solution of 60 µL of each UDE was added to 60 µL of DPPH (60 µM) in methanol. After mixing vigorously for 10 s, the mixture was transferred into a 100 µL Teflon capillary tube and the scavenging activity of each sample on DPPH radical was measured using a JES-FA ESR spectrometer (Jeol Ltd., Tokyo, Japan). A spin adduct was measured on an ESR spectrometer exactly after 2 min. Experimental conditions were as follows: central field, 3,475 G; modulation frequency, 100 kHz; modulation amplitude, 2 G; microwave power, 5 mW; gain, 6.3 x 105, and temperature, 298 °K.
Cell cultures and viabilities
BV2 murine microglia cells were cultured at 37 °C in 5 % CO2 in DMEM (Invitrogen, Carlsbad, CA, USA) supplemented with 5 % Fetal Bovine Serum (FBS, Hyclone, Logan, UT, USA) and antibiotics (Invitrogen). Cell viability was determined by 3-(4, 5- dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay as described previously [8]. In all experiments, cells were pre-treated with the indicated concentrations of UDE for 1 h before the addition of LPS (1 μg/mL, Sigma-Aldrich, St. Louis, MO, USA) in serum free DMEM. An equal volume of sterile water was added to all control treatments. Cells were incubated with various UDE concentrations for 24 h followed by MTT for 4 h, and then 100 μl of isopropanol (in 0.04 N-hydrochloric acid) was added to dissolve the formazan crystals. The absorbance was read at 570 nm using the Biochrom Anthos 2010 spectrophotometer (Salzburg, Austria). Cell viability was measured by the relative absorbances of the samples compared to the control.
BV2 murine microglial cells were grown in RPMI1640 (Invitrogen, Carlsbad, CA, USA), supplemented with 10 % FBS (Hyclone, Logan, UT, USA) containing 100 U/mL of penicillin (Invitrogen), and 100 μg/mL streptomycin (Invitrogen) in 37 °C in humidified atmosphere of 5 % CO2. In all experiments, cells were pre-treated with the indicated concentrations of UDE for 1 h before the addition of LPS (1 µg/mL, Sigma-Aldrich, St. Louis, MO, USA) in serum free RPMI1640. An equal volume of sterile water was added to all control treatments. Cell viability was determined by 3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay as described previously [8]. Cells were incubated with various concentrations of UDE for 24 h followed by MTT for 4 h, and then 100 μL of isopropanol (in 0.04 N-hydrochloric acid) was added to dissolve the formazan crystals. The absorbance was read at 570 nm using the spectrophotometer (Tecan, USA). Cell viability was calculated as relative absorbance compared to control.
Nitric oxide assay
The amount of stable nitrite, the end product of NO generation, inactivated microglia was determined by a colorimetric assay as previously described [8]. Each supernatant was mixed with the same volume of Griess reagent (1 % sulfanilamide in 5 % phosphoric acid and 0.1 % naphthyl ethylenediamine dihydrochloride in water). The absorbance of the mixture at 540 nm was determined with an ELISA reader (Bio-Tek Instrument, Winooski, VT, USA). Nitrite concentration was determined by extrapolation from a sodium nitrite standard curve.
Nuclear protein extraction and Western blot analysis
Cells were washed in cold PBS three times and lysed in a buffer containing 50 mM Tris-HCl, pH 7.4, 1 % (v/v) Tergitol- type NP-40 (NP-40), 0.25 % sodium deoxycholate, 150 mM NaCl, 1 mM EDTA, 25 mM NaF, 2 mM sodium orthovanadate (Na3VO4) and protease inhibitor cocktail (Complete MiniTM, Roche, Mannheim, Germany) at 4 °C. The lysate was clarified by centrifugation at 10,000 x g for 20 min at 4 °C to remove insoluble components. Cell lysates were normalized for protein content using bicinchoninic acid (BCA) reagent (Pierce, Rockford, IL, USA). Equal amounts of protein were loaded onto 10 % poly acryl amide gel electrophoresis (PAGE) and separated by standard sodium dodecyl sulphate (SDS)-PAGE procedure. Proteins were transferred to an NC membrane (S&S, Dassel, Germany) and blocked with 5 % non-fat dry milk in TBS. To detect protein expression, the blots were probed with the specific antibodies followed by the secondary antibodies coupled to horseradish peroxidase (Bio-Rad, Herculus, CA, USA) with β-actin as internal control. The immunoreactive proteins on the membrane were detected by chemiluminescence using the West-Save substrate (Lab-Frontier, Seoul, Korea) on X-ray film. The antibodies against inducible nitric oxide synthase (iNOS), cyclooxegenase (COX) -1, COX-2, nuclear factor kappa-B (NF-κB), I kappa B-alpha (IκB-α), on extracellular signal-regulated kinase (ERK 1/2) and β-actin were purchased from Cell (Beverly, MA, USA).
Pro-inflammatory cytokines assay
Murine BV2 microglia cells (1 x 105 cells/well) were cultured on 96 well plates and treated with the UDE at indicated concentrations for 1 h and stimulated with LPS (1 µg/mL). At 24 h post LPS treatment, the production of TNF-α and 1 h post IFN-ɣ treatment, the production of TNF-α were determined in cell supernatant using assay kits (BD Biosciences, San Jose, CA, USA) as per the manufacturer’s instructions. The pro-inflammatory cytokines assay was performed at room temperature and optical absorbance at 450 nm was measured using the ELISA reader.
Statistical analysis
All data are presented as mean ± S.E.M of at least three independent experiments. Statistical analyses were performed by SAS statistical software (SAS Institute, Cray, NC, USA) using one-way analysis of variance, followed by Dunnett's multiple range tests. P-values < 0.05 were considered statistically significant.
Results
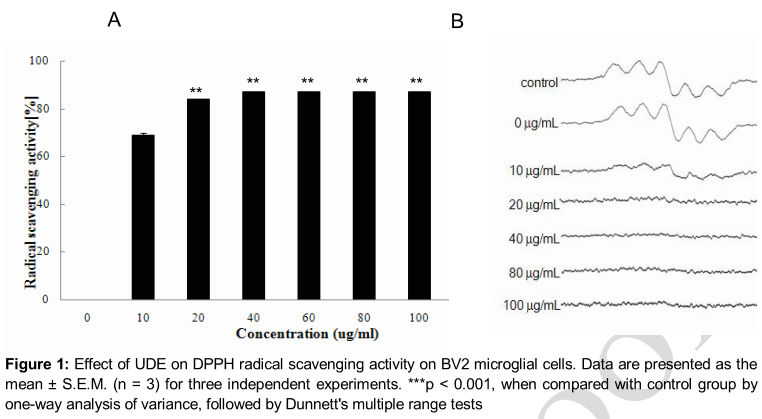
Effect of UDE on DPPH radical scavenging activity
As shown in A, UDE exhibited significant DPPH radical scavenging activity in a dose-dependent manner, showing a maximum effect at 100 μg/mL of concentration (p < 0.001). The ESR spectroscopy data at 10, 20, 40, 60, 80 and 100 μg/mL are shown in B. UDE significantly scavenged the DPPH free radicals.
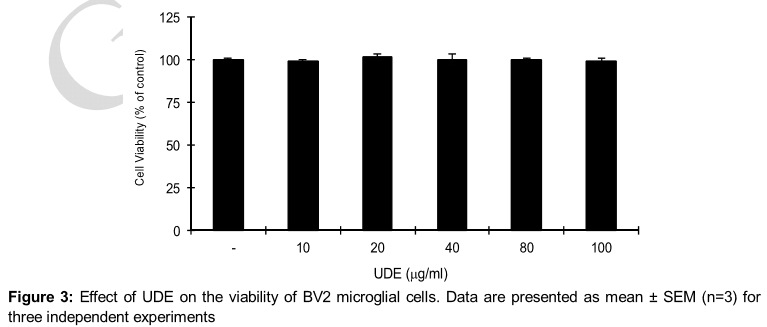
Effect of UDE on LPS-induced NO production
UDE treatment did not exhibit any significant cytotoxicity in BV2 murine microglial cells even at a high concentration (100 μg/ml) up to 24 h of incubation, and in all cases the viability was found above 97 % by MTT assay ().
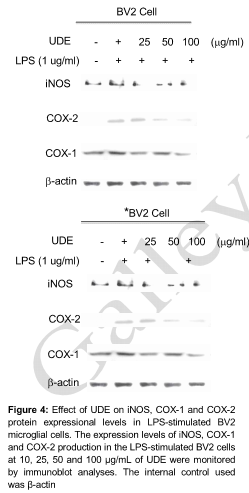
Effect of UDE on LPS-induced expression of iNOS and COX levels
UDE exhibited a broad spectrum of inhibitory effects on the expression of pro-inflammatory mediators. UDE reduced the LPS-stimulated increase of iNOS protein expression and inducible COX-2 in a concentration-dependent manner. This results clearly showed that UDE attenuated LPS-induced iNOS expression and downstream NO production. LPS strongly activates microglia and induces COX-2. Constitutive COX-1 protein expressional levels were uninterrupted ().
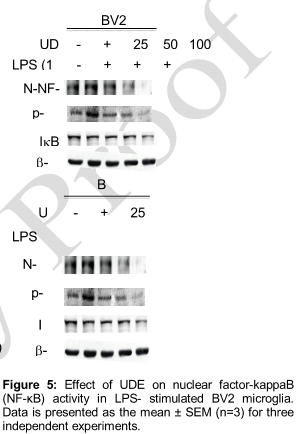
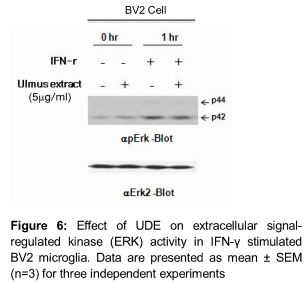
Effect of UDE on NF-κB and ERK levels
UDE inhibited the LPS-induced phosphorylation and degradation of IκB-α, and nuclear translocation of p65 NF-κB in a concentration- dependent manner (). In addition, UDE markedly inhibited the IFN-ɣ induced phosphorylation of ERK after 1h stimulation (). These results showed that UDE inhibited LPS-induced phosphorylation/degradation of IκBα and translocation of NF-κB/p65 sub-unit in a concentration-dependent manner.
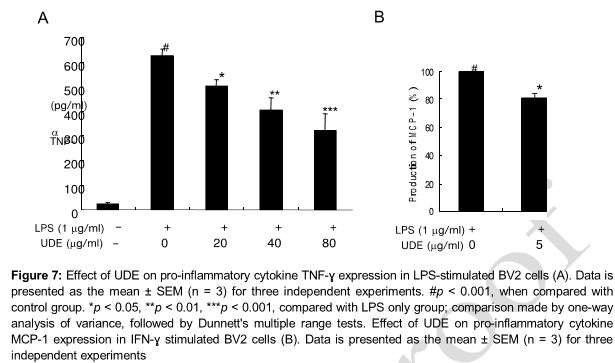
Effect of UDE on TNF-α production in LPS- stimulated BV2 cells
LPS stimulation of the BV2 murine microglial cells increased TNF-α expression. However, pretreatment with UDE decreased the LPS-induced TNF-α level in BV2 microglia (A). Pro-inflammatory cytokines such as TNF-α, IL-1β and IL-6 caused potent activation of iNOS gene expression in rodent glial cells and muscle cells [11]. The likelihood of the involvement of UDE in attenuating such factors is supported by this observations that pro-inflammatory mediator TNF-α produced by LPS treatment was suppressed by UDE in BV2 microglial cells indicating that UDE may be an effective anti-neuroinflammatory agent.
Effect of UDE on MCP-1 production in IFN-ɣ stimulated BV2 cells
Monocyte Chemo-attractant Protein-1 (MCP-1) is the first discovered and most extensively studied CC chemokine, and the number of studies on the etiologies of obesity and diabetes-related diseases has increased exponentially in the past two decades [12]. The roles of MCP-1 in the development of obesity, diabetes and cardiovascular diseases in metabolically healthy problems [12]. IFN-ɣ stimulation increased MCP-1 expression in BV- cells, while pretreatment with UDE decreased the LPS-induced MCP-1 levels in BV2 microglia (B).
Discussion
Inflammation is partially ascribed to release of toxic free radicals and ROS. DPPH radical scavenging assay is one of the widely used methods for screening the free radical scavenging activities of several agents in a relatively short period of time. In the present study, AEE significantly scavenged the DPPH free radicals. In this study a pro-inflammatory stimulus by LPS on BV2 cells resulted in excessive production of NO. NO production by LPS-activated cells was significantly inhibited by UDE in a concentration-dependent manner.
Excessive production of inflammatory mediators such as nitric oxide (NO), prostaglandin E2 (PGE2), and proinflammatory cytokines, including tumor necrosis factor-alpha (TNF-α) and interleukin-1beta (IL-1β) from activated microglia contributes to uncontrolled inflammation in neurodegenerative diseases. It seems possible that treatment with anti-inflammatory agents, including plants used in Oriental medicine, might delay the progression of neurodegeneration through the inhibition of microglial activation [9]. These results indicated that UDE significantly inhibited the excessive production of NO, in LPS-stimulated BV2 cells.
It was well known that COX-1 is constitutively expressed in many cell types and COX-2 is normally not present in most cells, but its expression is induced in response to inflammatory cytokines linked to pathological events [10]. Earlier literature revealed that increased expression of the COX-2 plays an important role in the inflammatory process and neuro-degenerative diseases [10]. Several reports indicate that iNOS and COX-2 are induced excessively in various types of central nervous injuries and diseases [10]. Attenuation of the induction of iNOS and COX-2 in activated microglia can prevent neuro-inflammation. Although this results point to a potential role of UDE in inhibiting iNOS and COX-2 production, the possibility remains that UDE may further suppress other microglia-derived pro-inflammatory factors upon LPS treatment in BV2 cells.
Pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 play central roles in inflammation. In particular, increased levels of brain TNF-α and IL-6 has been associated with severe cognitive impairments, neuronal damage and neuroinflammation [13]. Therefore, the effects of UDE on pro-inflammatory cytokine TNF-α production in LPS-stimulated BV2 cells were evaluated. LPS-stimulation increased the levels of TNF-α in BV2 cells. However, pretreatment with UDE suppressed the increased TNF-α production indicating that AEE may convincingly be an effective anti-inflammatory agent.
NF-κB, a mammalian transcription factor, activated by LPS, is known to control the expression of cell survival genes as well as proinflammatory enzymes and cytokines [14].
The molecular mechanisms that underlie UDE-mediated attenuation are related to the inhibition of the phosphorylation of three mitogen-activated protein kinases (MAPKs), extracellular signal-regulated kinases 1 and 2 (ERK1/2), p38 MAPK, and c-Jun N-terminal kinase (JNK), and the activation of nuclear factor-kappaB (NF-κB). Our result showed that AEE inhibited the LPS induced phosphorylation/degradation of IκB-α and translocation of NF-κB/p65 sub unit in a concentration-dependent manner. Considering the above data, we can conclude that NF-κB is a major target of UDE. However, the exact molecular target of UDE on NF-κB activation remains to be elucidated.
Conclusion
The findings of this study indicate that UDE inhibits anti-inflammatory responses via NF-κB signaling in LPS-stimulated BV2 murine cells. Further, the antioxidant activity of UDE may be partly involved in the observed effects.
Declarations
Acknowledgement
References
Archives


News Updates