Jun-Li Shao ![]() ,
Hai-Rong Liang,
Juan-Xiu Dai
,
Hai-Rong Liang,
Juan-Xiu Dai
For correspondence:- Jun-Li Shao Email: sjl@gdmc.edu.cn Tel:+8675922896573
Received: 1 October 2015 Accepted: 23 December 2015 Published: 29 January 2016
Citation: Shao J, Liang H, Dai J. Luteoloside inhibits proliferation of human chronic myeloid leukemia K562 cells by inducing G2/M phase cell cycle arrest and apoptosis. Trop J Pharm Res 2016; 15(1):39-45 doi: 10.4314/tjpr.v15i1.6
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose:To investigate the effects of luteoloside on the proliferation of human chronic myeloid leukemia K562 cells and whether luteoloside induces cell cycle arrest and apoptosis in K562 cells.
Methods:Luteoloside’s cytotoxicity was assessed using a cell counting kit. Cell cycle distribution was analysed by flow cytometry after propidium iodide (PI) staining. Cell apoptosis was assayed with apoptosis detection kit and Hoechst staining followed by observation under a fluorescence microscope. The ex
Results:Luteoloside inhibited the proliferation of K562 cells in a dose- and time- dependent manner (IC50 = 30.7 μM) with less toxicity in a normal human cell line (IC50 = 91.8 μM). Moreover, anti-proliferative effect of luteoloside was accompanied with G2/M phase arrestA288;p A308;0.05 or pA308;0.01A289;and apoptosisA288;p A308;0.01 or p A308;0.001A289;. Further studies revealed that the ex
Conclusion:These results suggest that the inhibitory effect of luteoloside on K562 cell proliferation is associated with inducing G2/M phase arrest and apoptosis, and that luteoloside is worth further studying for anticancer potential.
Introduction
Chronic myeloid leukemia (CML) is a malignant hematological disease characterized by the deregulated growth of myeloid leukemia cells in the bone marrow and their accumulation in the blood. The incidence of CML is 1 - 2 cases per 100,000 adults, and CML accounts for approximately 15 % of newly diagnosed cases of leukemia in adults [1]. Furthermore, CML is still one of the most difficult malignant hematological diseases to treat. The current commonly used chemotherapies are often accompanied with various side-effects, such as anemia, hemorrhage, infection, fever, enlargement of the liver, spleen and lymph nodes, and ostealgia [2]. Therefore, it is required to find new anticancer agents with increased efficacy and decreased toxicity.
Plant-derived compounds have attracted considerable attention in cancer chemoprevention and treatment because of their high efficiency and safety [3]. Luteoloside, a natural flavone subclass derived from plants, has exhibited a series of biological activities, such as potential antibacterial and antifungal [4], free radical scavenging [5], anti-oxidative capabilities [6,7], protection effects against doxorubicin-induced cardiotoxicity [8], and anticarcinogenic potential against colon carcinogenesis [5] and hepatocellular carcinoma [9,10], etc. However, the effects of luteoloside on blood malignancy have not been reported.
Here we carried out a study on the cytotoxicity of luteoloside against human CML K562 cells. K562 cells were derived from CML patient carrying a BCR-ABL mutation. The mutation gives poor response to treatment by deregulated apoptosis, proliferation and differentiation [11]. Since an important mechanism for anticancer agents is to trigger apoptosis [12] and cell cycle arrest [13] in cancer cells while disturbing their proliferation, we evaluated the effects of luteoloside on cell cycle and apoptosis together with cell growth of K562 cells. To further explain the underlying mechanism of luteoloside effects, the expression of cell cycle- and apoptosis-related regulatory proteins were assessed in K562 cells treated with luteoloside.
Methods
Cell culture, antibodies and chemicals
Leukemia cell K562 and human normal cell HUVEC12 were cultured in RPMI-1640 and DMEM media respectively. All of them were cultured in the media supplemented with 10 % fetal bovine serum at 37 oC in a humidified 5 % CO2 atmosphere. Antibodies against cyclinB1, Bax and Bcl-2 were purchased from Cell Signaling (Boston, USA). NAPDH antibody and horseradish peroxidase-conjugated secondary antibody were purchased from Beyotime Biotechnology (Beijing, China). Cell Counting Kit-8 (CCK-8) and annexinV-FITC apoptosis detection kit were purchased from DoJindo (Kumamoto, Japan).
RNase A, propidium iodide (PI), dimethyl sulfoxide (DMSO) and Hoechst 33342 were purchased from Sigma-Aldrich (St. Louis, USA). RIPA lysis buffer and BCA protein assay kit were purchased from CWBIO Biotechnology (Beijing, China). Luteoloside was purchased from Shanghai Winherb Medical Technology Co., Ltd. (Shanghai, China). Luteoloside was dissolved in DMSO at 25, 50, 100 and 200 mM as stock solutions and stored at -20 oC, and diluted 1000 times with media as work solution when used.
Cell growth inhibition assay
HUVEC12 cells were seeded into 96 well microtiter plate (3 × 103 cells per well), and allowed to attach for 16h. Then media were exchanged with fresh media containing luteoloside at the indicated concentrations. K562 cells were seeded into 96 well microtiter plate (3 × 103 cells per well) together with various concentrations of luteoloside. Cells treated with 0.1 % DMSO were used as controls. After treatment for 48 and 72 h, the cytotoxicities of luteoloside against K562 and HUVEC12 cells were assessed using CCK-8 assay. Briefly, CCK-8 reagent (10 μL) was added to each well and the cells were incubated for 2 h at 37 °C. Absorbance was then analysed on a microplate reader (BioTek Synergy2) at 450 nm. All experiments were done in triplicate.
Cell cycle analysis
K562 cells were treated with luteoloside for 48h at concentrations of 50 and 200 μM or for 24 and 48 h at 50 μM. At the end of the treatment, the cells were collected by centrifugation (1000 rpm, 5 min), washed twice with ice-cold PBS, and fixed in 75 % ethanol at -20 oC overnight. Then, cells were washed twice with ice-cold PBS containing 0.2 % TritonX-100, and stained with DNA-staining solution (50 μg/mL PI and 100 μg/mL RNase A in PBS) in the dark for 30 min at room temperature. Finally, cells were analysed by a flow cytometer (Becton Dickinson, CA, USA). All experiments were in duplicate and repeated three times.
Detection of apoptosis
K562 cells were treated with luteoloside at different concentrations (50 and 200 μM) for 48 or 72 h. Then, apoptosis was assayed with annexinV-FITC apoptosis detection kit following the manufacturer’s instructions. Briefly, cells were harvested by centrifugation, washed twice in PBS and re-suspended in binding buffer. AnnexinV-FITC and PI were then added to the cell suspension. After incubation for 15 min in the dark at room temperature, cells were diluted with binding buffer again and the ratio of apoptotic cells was measured within 1 h with a flow cytometer. All experiments were measured in duplicate and repeated three times.
Hoechst 33342 staining was performed to detect the alterations of nuclear morphology of K562 cells after luteoloside treatment. Hoechst 33342 was added to the treated cells at a final concentration of 10 μg/mL, and the cells were incubated for another 10 min at 37 oC. The stained cells were then observed to examine the degree of nucleus condensation under a fluorescence microscope equipped with standard excitation filters (Nikon, Japan).
Western blot analysis
Cells treated with luteoloside at the indicated concentrations and for different incubation time were harvested, washed twice with ice-cold PBS and lysed in ice-cold RIPA buffer for 10 min on ice. The lysates were centrifuged at 12,000 rpm for 10 min at 4 oC, and supernatants were collected. Protein concentrations were determined with BCA protein assay kit. Equal amounts of protein sample (30 μg) were fractionated on SDS-PAGE, and electro-transferred onto PVDF membranes (Millipore). After blocking with 5 % (w/v) non-fat dry milk in PBS for 1 h at room temperature, the membranes were incubated overnight at 4 °C with the specific primary antibodies with a 1:1000 dilution. The membranes were washed and then further incubated with horseradish peroxidase-conjugated secondary antibody (1:1000) for 2 h at room temperature. After the membranes were washed again, the protein bands were visualized using a chemiluminescence imaging method (Pierce ECL Western Blot Substrate). All blots were repeated three times.
Statistical analysis
Data are presented as mean ± standard deviation (SD). Significant differences between treated and untreated (control) cells were identified by Student’s t test. P < 0.05 was considered statistically significant. IC50 (half maximal inhibitory concentration) values were calculated by regression analysis.
Results
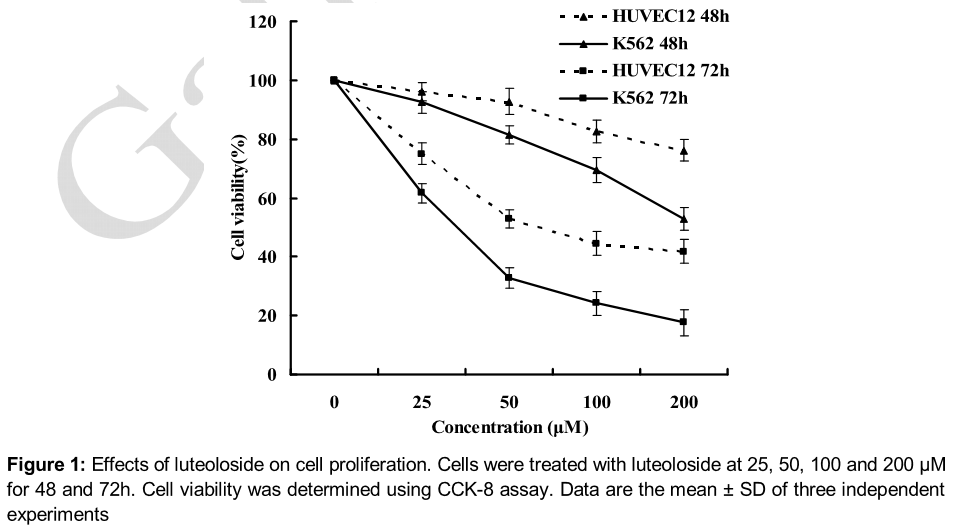
Luteoloside inhibited the proliferation of K562 cells
Anti-tumor drugs are often screened according to their cytotoxic effects on cell proliferation or viability. Therefore, we first investigated the effects of luteoloside on the proliferation of K562 cells exposed to luteoloside at various concentrations and for different time interval. As shown in , luteoloside significantly inhibited the proliferation of K562 cells in a concentration- and time-dependent manner. When K562 cells were treated for 72h, the IC50 value was 30.7 μM. Moreover, there were smaller cytotoxic effects of luteoloside on the normal cell line HUVEC12 at the tested concentrations, with an IC50 value of 91.8 μM for 72 h treatment.
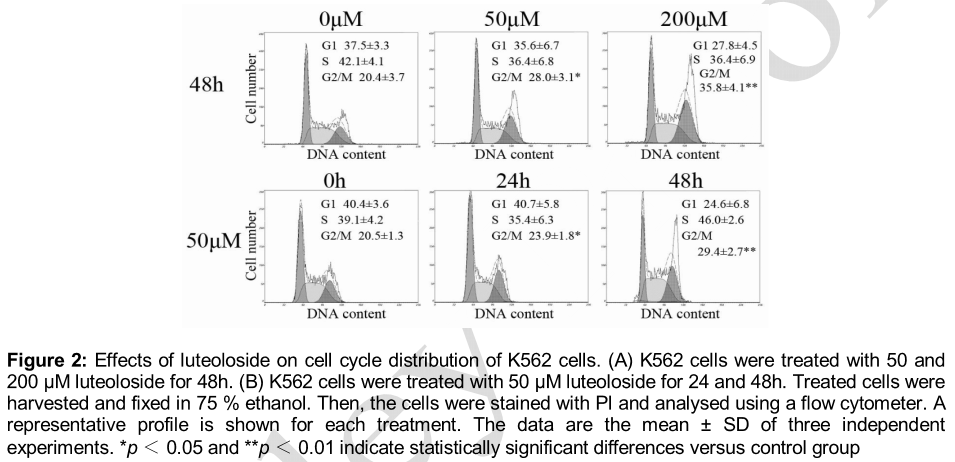
Luteoloside induced cell cycle arrest at G2/M phase
To investigate the mechanism behind the inhibition of cell growth by luteoloside, cell cycle analysis was conducted by flow cytometry. As shown in A, the percentage of G2/M phase increased as the concentration of luteoloside increased in the 48 h treatment, which was 28.0 (p < 0.05) and 35.8 (p < 0.01) at 50 and 200 μM, respectively, while that of the untreated cells was only 20.4.
B shows that the percentage of G2/M phase increased with prolonged incubation time at 50 μM, which was 23.9 (p < 0.05) and 29.4 (p < 0.01) for 24 and 48 h treatment respectively, while that of the untreated cells was only 20.5. These results demonstrate that luteoloside induces G2/M phase cell cycle arrest in K562 cells in a concentration- and time-dependent manner.
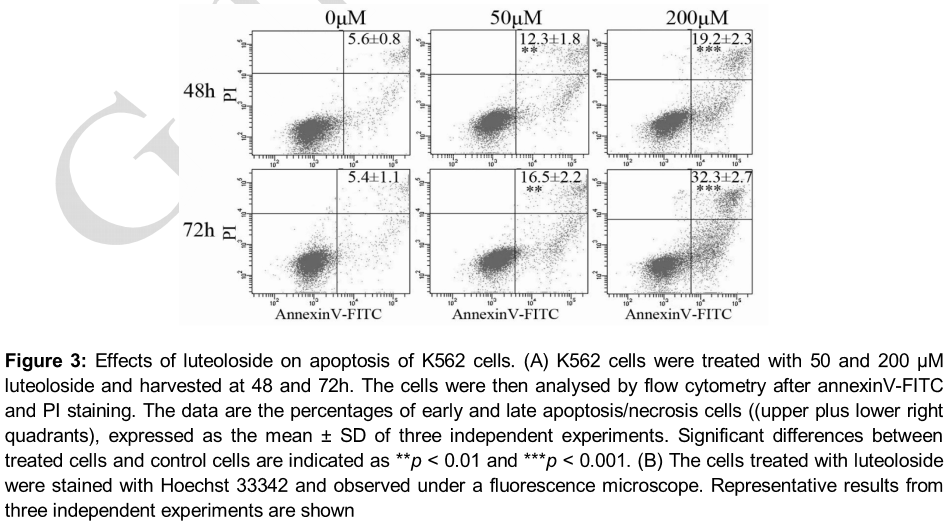
Luteoloside-induced apoptosis
Using flow cytometry after annexinV-FITC and PI staining, we evaluated whether luteoloside induced apoptosis in K562 cells. As it can be observed (A), luteoloside increased the number of total apoptotic cells 2.2-fold at 50 μM and 3.4-fold at 200 μM for 48h treatment and 3.1-fold at 50 μM and 6.0-fold at 200 μM for 72h treatment. Therefore, apoptotic effects of luteoloside on K562 cells were in a dose- and time-dependent manner. Since cell apoptosis is accompanied with alterations of nuclear morphology, Hoechst 33342 staining was performed to detect alterations of nuclear morphology of K562 cells treated with luteoloside. As shown in B, the nuclei of the control cells were uniformly stained, while the nuclei of the treated cells showed the typical characteristics of apoptosis, including condensed chromatin and punctate apoptotic bodies.
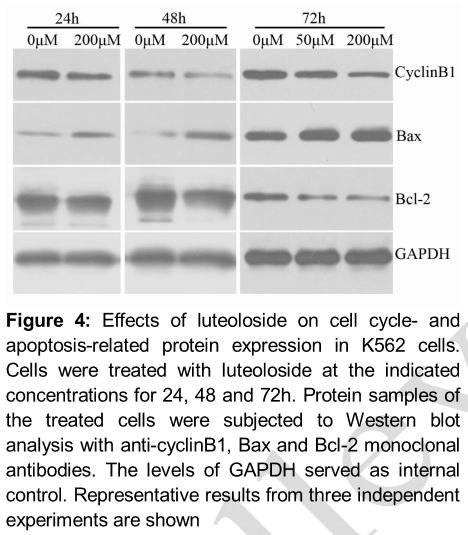
Luteoloside decreased cyclinB1 and Bcl-2 expression and increased Bax expression
Expression of the intracellular proteins related to G2/M phase arrest and apoptosis, such as cyclinB1, Bax and Bcl-2, was investigated to understand the mechanisms by which luteoloside inhibited K562 cells. As shown in , the expression of cyclinB1 decreased in response to luteoloside treatment. The expression of the pro-apoptotic protein Bax was upregulated markedly, while the expression of the anti-apoptotic protein Bcl-2 was suppressed by luteoloside at the indicated concentrations and for the incubation time.
Discussion
Since rapid growth due to deregulated cell cycle and reduced apoptosis is the important property of cancer cells, inhibiting proliferation by arresting cell cycle progression and promoting apoptosis has been an important strategy for anticancer drugs and chemo-preventive agents [12,15-17]. In this study, it is revealed that luteoloside can induce cell cycle arrest and apoptosis concomitantly with growth suppression of K562 cells. Thus, luteoloside has exhibited its anti-cancer effects not only on solid tumors [5,9,10] but also on blood malignancy.
Considering the side-effects of the current chemotherapy drugs, it is required to conduct research on agents with high efficiency and low toxicity. In this study, luteoloside showed high cytotoxicity on K562 cells in a dose- and time-dependent manner with an IC50 of value of 30.7 μM for 72 h treatment. In addition, toxicity evaluation on the normal cell HUVEC12 revealed only minor decrease in cell viability at the tested concentrations with an IC50 of 91.8 μM for 72 h treatment. In contrast, luteoloside suppressed proliferation of several hepatocellular carcinoma cells [10], colon carcinoma cell COLO 320 DM and normal cell VERO [5] also in a dose- and time-dependent manner with IC50 of about 75, 112.4 and 724 μM, respectively, for 72 h incubation. Therefore, the cytotoxic effects of luteoloside on K562 are in high efficiency and specific to some extent.
Reproducing healthy cells is strictly regulated due to rigorously and accurately controlled cell cycle. Control of the cell cycle is driven by the cyclin-dependent kinases (CDKs). Cells cannot enter S phase or M phase without CDK activation. In order to become catalytically active, CDKs need to bind to a cyclin subunit that acts as an activator [15]. CyclinB1 is one of main cyclins involved in the regulation of the transition from G2 to M phase during the cell cycle. Modification subsequent to CDK1 binding to cyclinB1 causes activation of the CDK1/cyclinB1 complex and drives the cell into mitosis [18]. CylclinB1 is synthesized initially in late G1 phase and its content reaches a maximum in G2 phase. It was reported that some natural or chemical compounds that cause G2/M phase arrest involved the down-regulation of cyclinB1 [19,20]. In the present study, luteoloside treatment increased the cell number at G2/M phase, as confirmed by flow cytometry. Western blot analysis further revealed the decreased expression of cyclinB1. These results indicate that down regulation of cyclinB1 may contribute to luteoloside-induced G2/M arrest in K562 cells.
Apoptosis, also known as programmed cell death, plays an important role in the regulation of development and homeostasis [21,22]. Cells undergoing apoptosis always show a series of characteristics including cell shrinkage, membrane blebbing, chromatin condensation, DNA fragmentation and, eventually, cellular breakdown into apoptotic bodies [23].
Apoptosis occurs through two main pathways: the extrinsic pathway, known as death receptor pathway, and intrinsic pathway, known as mitochondria pathway [24]. Mitochondria play a central role as the key control point in the mitochondria pathway. Bcl-2 family proteins, outer mitochondrial membrane proteins, have been well known as crucial regulators in the mitochondria pathway. Based on function, they are divided into two subclasses, pro-apoptotic Bcl-2 family proteins (Bax, Bak and Bad, etc.) and anti-apoptotic Bcl-2 family proteins (Bcl-2, Bcl-xL and Mcl-1, etc.). Whether mitochondria pathway is activated depends on the balance between pro- and anti-apoptotic Bcl-2 family proteins. High expression of pro-apoptotic Bcl-2 family proteins can enhance the permeability of outer mitochondrial membrane and the consequent translocation of cytochrome c from the mitochondria to the cytoplasm, which provokes the downstream apoptotic events, while the anti-apoptotic Bcl-2 family proteins oppose the pro-apoptotic Bcl-2 family proteins [25-27]. Increasing Bax/Bcl-2 ratio often accompanies phytochemical-induced apoptosis through mitochondrial pathway in cancer cells [28-30].
In this study, luteoloside treatment triggered cell death of K562 cells by inducing apoptosis, which was confirmed by an increase in the percentage of total apoptotic cell population and the condensation or fragmentation of chromatin/ nuclei after luteoloside treatment. Further study displayed that the pro-apoptotic protein Bax increased and the anti-apoptotic protein Bcl-2 decreased after luteoloside treatment. These results indicate that luteoloside-induced apoptosis is mediated by Bax and Bcl-2 through the mitochondrial pathway. However, it cannot be excluded that the extrinsic apoptotic pathway is involved in the luteoloside-induced apoptosis based on the present results.
It should be noted that future studies are required to investigate more molecular events responsible for luteoloside-induced cell cycle arrest and apoptosis.
Conclusion
Despite its preliminary characteristics, this study indicates that luteoloside has antitumor potentials due to its growth inhibitory effect, G2/M phase arrest and inducement of apoptosis. Furthermore, decreased cyclinB1 expression may contribute to G2/M phase cell cycle arrest. Besides, up-regulation of pro-apoptotic molecule Bax and down-regulation of anti-apoptotic molecule Bcl-2, demonstrate that intrinsic apoptotic pathway is one of the pathways through which luteoloside-induced apoptosis occurs.
Declarations
Acknowledgement
References
Archives


News Updates