Xiang-Xiu Sun1,
Qing-Shan Ma2,
Tian-Feng Liu1,
Ying Chen1,
Yan Dong1,
Lin-Lin Zhang1 ![]()
For correspondence:- Lin-Lin Zhang Email: zhanglingling49@gmail.com Tel:+865398216079
Received: 4 August 2015 Accepted: 1 December 2015 Published: 29 January 2016
Citation:
Sun X, Ma Q, Liu T, Chen Y, Dong Y, Zhang L.
Aldehyde dehydrogenase 1 and Raf Kinase inhibitor protein ex
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To determine the differences in response following the use of aldehyde dehydrogenase 1 (ALDH1) and Raf kinase inhibitor protein (RKIP) as cervical cancer stem cell markers.
Methods: To evaluate the cancer stem cell markers, a mouse model with low and high grade cervical cancer was developed and studied by histological examination. Immunohistochemical and Western blotting techniques were employed to study the ex
Results: Histological data helped to differentiate low from high grade cervical cancer. The results from immunohistochemistry show increased pattern of Sox2 ex
Conclusion: ALDH1 and RKIP marker in association correlation with Sox2 aids in defining the proliferative ability of cervical cancer stem cells and also to validate them in initial and advanced stages of cervical cancer.
Introduction
On a global level, cervical cancer is reported to be the fourth most common cancer among women population [1]. Yearly, almost 470,000 new cases are diagnosed with a malignant form of cervical cancer and 233,000 death cases are reported every year [2]. Chemotherapy, radiotherapy and surgery are handful to improve the survival of patients with cervical carcinoma [3], but in approximately 35 % of cases, after initial improvement, the disease starts to worsen within the time range of three years [4]. The recurrence of the disease implies that the resistance of cervical cancer cells or cancer stem cells that still remain after treatment gets triggered before their re-emergence. Cancer stem cells possess the ability of self-renewal and differentiation to form matured cancer cells that are linked with tumour initiation, metastasis and recurrence [5]. The role of cancer stem cells was first identified in leukemias and later, were identified in solid tumours like brain [6], colon [7] and also in cervical cancers [8].
The field research area of cancer stem cells is a newly developing field and identifying these cells is tricky at present. Most of the normal stem cell markers cannot effectively guide in the specific identification of cancer stem cells because it emerged from a specific mutation that takes place in stem cell population. Comparing the novel markers that show the under-expression and up-regulation in cancerous activity, helps to refine and identify the proliferative cancer stem cells. Aldehyde dehydrogenases (ALDH) are NAD(P)+ dependent detoxifying enzymes that determines cell survival ability was initially identified established as a stem cell marker in haematopoietic stem cells [9]. A high expression of ALDH can also used to distinguish neural stem cells [10]. The ALDH activity can also be a useful cancer stem cell marker because it determines metastasis and cell survival [11]. It is successfully used as a stem cell marker for many cancer types including cervical cancer [12]. The current study defines the role of ALDH as a cancer stem cell marker that vary among different cancer types and also within the same tumour cell lines [13]. The Raf kinase inhibitor protein (RKIP) was identified as a metastasis suppressor in many tumour types like in prostate cancer [14], colorectal cancer [15] and in cervical cancer [16]. But these markers suffer from poor prognosis in cervical cancer, as well as in other cancer types [17,18]. In this paper, we have selected a pair of over-expression and under-expression markers namely ALDH and RKIP, to understand the proliferative nature of cancer stem cells in a cervical cancer mice model so that a precise identification is achieved.
Methods
Animals
The animals were handled following the guidelines for using experimental animals [19]. For experimental purpose, the female strain of nude athymic mice (BALB/c-nu/nu), aged 8 weeks was uniformly chosen. The HeLa cells grown in a serum free medium were dissociated to obtain individual cells. The cells were then resuspended in 100 μl of FBS and growth factor-free DMEM at a cell density of 1 × 105 cells [8]. The obtained cells were then injected subcutaneously into the left and right flanks [8]. The injected athymic mice were monitored and provided water, feed and maintained in dust free laboratory condition [20] for eight weeks. The tumour samples were collected in initial development stage of the tumour initiation (on the 4th week) and in the aggressive stage of the tumour condition (on the 8th week) by scarifying the mice using standard procedure [21].
Immunohistochemistry
For performing immunohistochemistry, the tissue sections were initially fixed in 10 % formalin solution, and after dehydration and clearing, the tissues were paraffin embedded. The tissue sections were then subjected to microtome sectioning of size 6 µm. The sections were then carefully placed on glass slides and were de-paraffinized and then rehydrated. The endogenous peroxidase activity was blocked by immersing the sections in freshly prepared 10 % H2O2 and 10 % Methanol with 1X PBS for 20 mins. Trypsin treatment (0.1 % trypsin in 0.1 % CaCl2) was maintained for 10 min to cleave the protein cross link, so as to assess the antigen and epitope. The non-specific antigens were blocked using 4 % of BSA and then were incubated with suitable primary antibody (Anti-ALDH1 antibody, Abcam (EP1933Y); Anti-Sox2 antibody, Abcam (ab92494); anti-RKIP Antibody, Santa cruz (SC-28837) overnight at 4 oC. After incubation, the sections were thoroughly washed with 1X PBS and latter incubated with a suitable secondary antibody for 30 min at room temperature. After washing the non-specific binding, the sections were stained with DAB (diaminobenzidine) Kit.
Western blot analysis
The samples from normal tissue, initial stage and samples from advanced stages of cervical cancer were dissected and the protein samples were prepared from the cell lysate. The cellular proteins were resolved in 12 % SDS-PAGE gel by following the protocol as described [22]. The protein in the gel was then transferred to the PVDF membrane. After blocking the membrane overnight at 4 ºC, they were incubated with an appropriate dilution of the primary antibody (Anti-ALDH1 antibody, Abcam (EP1933Y); Anti-Sox2 antibody, Abcam (ab92494); anti-RKIP Antibody, Santa cruz (sc-28837) according to the manufacturer’s instruction. The membranes were then incubated with the suitable secondary antibody of dilution concentration (1:3000). After following washing, the membrane was developed to obtain the signal.
Statistical analysis
The results are presented as mean ± standard deviation (SD) and were analysed by analysis of variance (ANOVA). The software used for the analysis was Microsoft Office Excel 2007. P < 0.05 was considered statistically significant.
Results
Mice model to study cervical cancer
Studies carried out with the cell lines of cervical cancer to find out new markers of cancer stem cells varies in result even within a particular tumour cell line and it requires different understanding [13]. Therefore, studying and assessing the cancer stem cells markers in an identical animal model was more valuable. Cervical cancer was developed in the in vivo model of mice as described in the materials and methods.
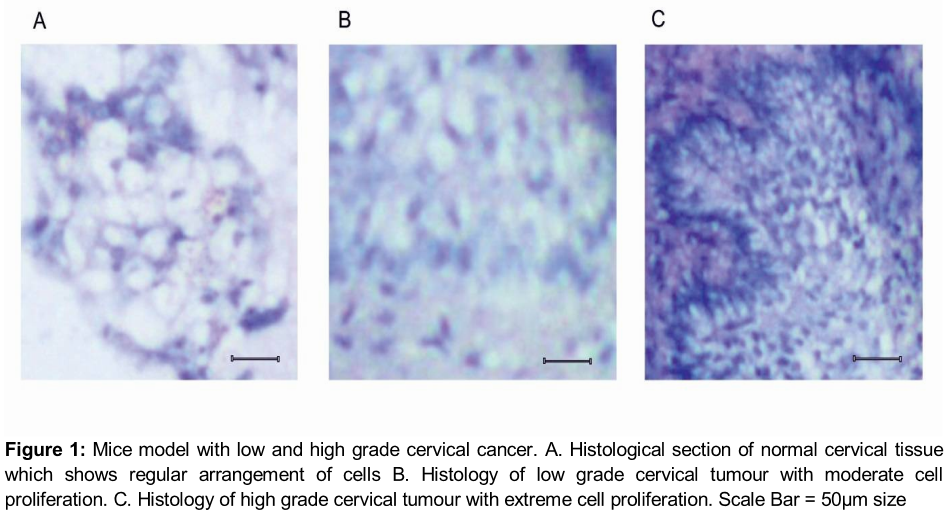
After injection with HeLa cells, the mice developed the initial stage of cervical cancer on the 4th week and more advanced stages of the tumour on the 8th week of post injection as visualized in the histological sections (). The histology of the normal cervical tissue sections shows the regular arrangement of a cellular pattern with less cell density (A). As the tumour starts to developed, the proliferative cells starts to increase during the initial stage on the 4th week (B) and reach its maximum during the advance stage of cervical cancer (C).
Comparing Sox 2 expression with ALDH and RKIP
The Sox2 expression was tightly correlated linked with the progression of many human malignancies [23] and it was effectively used in breast cancer and glioblastoma as a cancer stem cell marker [24,25]. The correlation association between Sox2 expression with cervical cancer stem cell population is also well documented [23]. Another interesting correlation is that the depletion in Sox2 expression is linked with the reduction of the transformational properties of the cell [26]. In this study, we used the Sox2 as a standard maker to determine the role of ALDH and RKIP in determining the proliferation ability of cervical cancer stem cells.
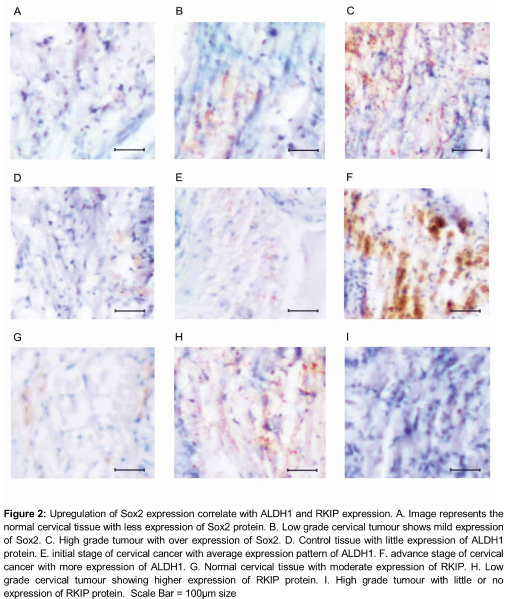
Immunohistochemical analysis of normal cervical tissue shows limited positive cells for Sox 2 expression (A). As the tumour progressed, the population of Sox2 positive cells shows concurrent increase in number by the 4th week of the initial stage of cervical cancer (B) and their expression reached its maximum on the 8th week of the advanced stage of the cervical cancer (p < 0.05) (C). Similarly, the ALDH1 expression also shows high-regulated expression as the tumour progressed (D-F).
The proposition of ALDH1 positive cells were limited in normal tissue (D) and their expression starts to increase in the primary stage of cancer (E).
As the tumour progressed to a severe stage on the 8th week, the ALDH1 expression shows the maximum extent (p < 0.05). The study was further extended to find out the positive regulation of RKIP in tumour regulation. In the control tissue, the RKIP directs higher expression (G) and acts as positive regulator that keeps check on the tumour progression. Interestingly, in the initial stage of the tumour, their expression gets up-regulated (H) but in the advanced stage of the tumour development, their expression gets down-regulated as expected (p < 0.05) (I).
Western blotting results
For better understanding of the correlation that the Sox2 expression has with ALDH1 and RKIP, the immunohistochemical data was further validated using western blotting analysis.
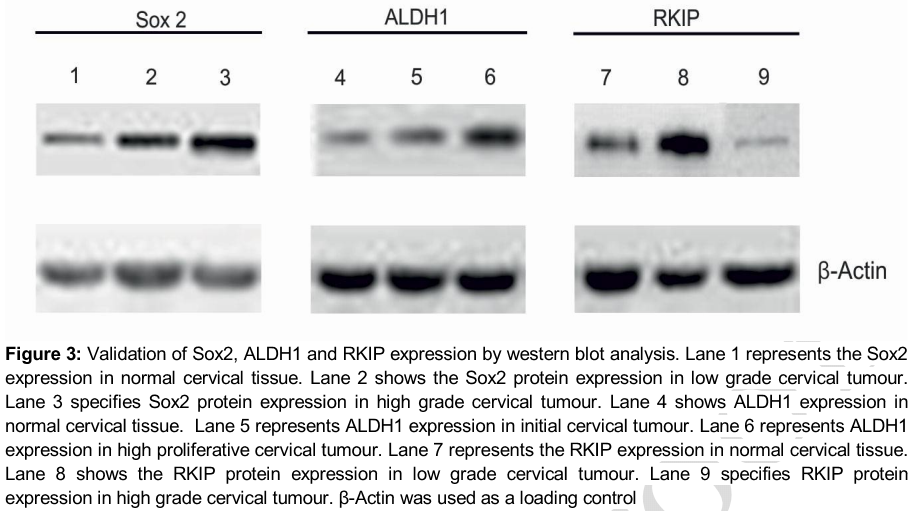
The results with western blotting analysis demonstrate shows that the protein expression profile of Sox2 dramatically increased as the tumour progressed as shown in first three lanes of . Similarly, increased the up regulated expression of ALDH1 with significant enhancement of ALDH1 expression in the advanced stage of the cervical cancer was noted as shown in lane four to six of . But when compared with the control tissue, we observed that the RKIP expression showed elevated expression profile in the initial stage of the tumour and down-regulated latter in the advance stage of the tumour progression development as shown in lane seven to nine of .
Discussion
The connections between stem cell markers with different pathological conditions of cancer were still poorly investigated. Recently it was revealed that new stem cell markers emerged while studying the process of tumour initiation and progression [27]. The present study was carried out with a well-documented stemness associated marker named Sox2 [23]. Using this standard marker, we evaluated assess the expression of ALDH1 and RKIP which are less sensitive markers, in different in vivo pathological conditions.
In recent studies of prostate cancer, the expression of ALDH1 shows relatively lower expression instead of high expression, but the result with other isoforms of ALDH remains the same [28]. Thus the results with ALDH1 are needed to be validated further; especially in their in vivo condition. In our study, the mice model responded well in the tumour formation and it formed low grade as well as higher grade tumours (B,C) that helps to study the function of different proteins in tumour progression. The results obtained with an immunohistochemical against ALDH1, shows extensively high label proliferative cells in the advance stage of cervical cancer (F) and the obtained result was consistent reliable with previously already reported data [12]. The higher the population of ALDH1 positive cells, the more revealed is their role in tumourigenicity and metastasis and their correlation is parallel with the cervical cancer stem cells that are identified using Sox2 (A-C). The study of RKIP expression in various grade of tumour, reveals that in the initial stages of the tumour, the RKIP has a control over tumour development with higher expression (H), but at the higher grade of the tumour their control over the tumour was relaxed with lower expression (I). Comparing the results along with Sox2, the higher expression upregulated profile of ALDH1 and lower expression of RKIP might also be involved in the pathogenesis of cervical cancer.
Conclusion
The expression pattern of ALDH1 and RKIP are tightly regulated in cancer stem cells in advanced stages of cervical cancer but they show disordered expression in initial stages of cervical cancer.
Declarations
Acknowledgement
References
Archives


News Updates