Xiao-xian Lu,
Xiao-xia Ji,
Jie Bao,
Qian-qian Li,
Dan-dan Ji,
Liang Luo ![]()
For correspondence:- Liang Luo Email: doctorluoliang@126.com Tel:+8651066681023
Received: 10 November 2015 Accepted: 8 June 2016 Published: 31 July 2016
Citation: Lu X, Ji X, Bao J, Li Q, Ji D, Luo L. Inhibition of proliferation, migration and invasion of human non-small cell lung cancer cell line A549 by phlomisoside F from Phlomis younghusbandii Mukerjee. Trop J Pharm Res 2016; 15(7):1413-1421 doi: 10.4314/tjpr.v15i7.9
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To determine the effect of phlomisoside F (PMF) on the proliferation, migration and invasion of human non-small cell lung cancer cell line A549 and explore the possible mechanisms.
Methods: The anti-proliferative effect of PMF on A549 cells was determined by CCK-8. Subsequently, migration and invasion were evaluated by Transwell and Transwell with matrigel assays, respectively. Furthermore, cell cycle and apoptosis were assessed by flow cytometry, while the mechanisms of action were determined by Western blotting.
Results: PMF exhibited significant anti-proliferative effect on A549 cells in concentration-dependent and time-dependent manners, with half maximal inhibitory concentration (IC50) of 54.51 µM. Treatment with PMF (10, 20 and 40 µM) for 48 h resulted in significantly decreased migration and invasion in A549 cells. In addition, PMF at concentrations of 25, 50 and 75 µM induced cell cycle arrest in G0/G1phase and enhanced cell apoptosis in A549 cells. Furthermore, caspase-3, caspase-9 and Bax protein ex
Conclusion: PMF suppresses A549 cell growth, migration and invasion. The mechanism may be related to the induction of mitochondria-mediated apoptosis pathway via regulation of caspase-3, caspase-9, Bcl-2 and Bax ex
Introduction
Lung cancer is the most common cancer in males with increasing incidence and mortality, and is the leading cause of cancer-related deaths in China [1]. Scientific data has clearly shown that the dominant risk factor of lung cancer is tobacco exposure and approximately 60 - 70 % of lung cancer patients are ever-smokers in China [2]. In addition, the most common type of lung cancer is non-small cell lung cancer (NSCLC) which has high rates of unknown disease stage at diagnosis [3]. Although researchers have made progress in the treatment of lung cancer, current treatment modalities are still inadequate and there is no cure. Therefore, an effective therapeutic method is still urgently needed to reduce the incidence and improve the survival of Chinese lung cancer patients.
Phlomis younghusbandii Mukerjee (Lamiaceae) is a perennial herbal plant, which is often found in the grassland of hillsides, meadows and river-beaches in Qinghai-Tibet Plateau [4]. Importantly, the aerial part of this plant has been used as a kind of Tibetan conventional medicine for thousands of years in China [5]. P. younghusbandii has been widely used in clinical treatment and has various pharmacological activities, such as treating lung disease, colds and coughs, bronchitis and throat disease [5,6]. However, research and study on modern pharmacology of P. younghusbandii is not common. Over the years, several studies demonstrated that the root extracts of P. younghusbandii had anti-bacterial, analgesic and anti-inflammatory activities [7-9]. But whether the extracts or chemical components from Phlomis younghusbandii have anti-cancer activity has not been reported up to now.
Phlomisoside F (PMF), a furanolabdane diterpene glycoside, is from the ethyl acetate extract of P. younghusbandii root, which has been reported in previous studies [10,11]. Previous data indicated that PMF has significant anti-inflammatory activity in vitro and in vivo [10]. In this study, PMF was isolated and identified from P. younghusbandii. Furthermore, the anti-lung cancer activity of PMF was investigated and the possible mechanism was explored in A549 cells, which may provide a scientific basis for its use in the traditional treatment of lung cancer.
Methods
Preparation of pure compounds
P. younghusbandii was procured from Hehuachi Market of Traditional Chinese Herbs in 2014 and identified by a taxonomist at the Department of Traditional Chinese Medicine of our hospital. A voucher specimen of P. younghusbandii (201408-LL) was kept in our laboratory for future reference. PMF was isolated from the dry root of P. younghusbandii and characterized by a previously described method [11]. In brief, the dry root of P. younghusbandii (10 kg) was pulverized to prepare the extract. After reflux extraction with ethanol two times for 2 h each time, the extract was filtered and then the solvent was evaporated to get the ethanol extract of P. younghusbandii root under reduced pressure in a rotary evaporator. Further, the crude ethanol extract was dissolved in distilled water and then the suspension was successively treated with petroleum ether, ethyl acetate and n-butanol. Thereafter, five fractions (I – V) were obtained by using silica gel (200 – 300 mesh) column chromatography and eluting with trichloromethane – methyl alcohol (8:1). Finally, the compound was isolated from fraction III by using repeated silica gel column chromatography (200 – 300 mesh) and Sephadex LH - 20 chromatography. Additionally, the isolated compound, phlomisoside F, was identified based on the 1H-NMR and 13C-NMR data, which was consistent with previous data [11]. The chemical structure of PMF is shown in .
Cell culture
Human non-small cell lung cancer cell line A549 was purchased from Shanghai cell bank of Chinese Academy of Sciences (Shanghai, China) and cultured in RPMI – 1640 medium (Hyclone, Logan, UT) supplemented with 10 % fetal bovine serum (FBS; Gibco, Grand Island, USA) and antibiotics (100 U/mL penicillin G and 100 mg/mL of streptomycin) at 37 °C in a 5 % CO2 humidified atmosphere.
Cell proliferation assay by CCK-8
Cellular proliferation was evaluated by Cell Counting Kit-8 (Beyotime, Jiangsu, China) following the manufacturer’s protocol. In brief, 2 × 103 cells per well were seeded in 96-well plates and incubated with either vehicle (DMSO) or increasing concentrations of PMF. Cell proliferation was measured 48 h later and the absorbance was measured using a 96-well plate reader (Bio-Rad Laboratories, Shanghai, USA) at 450 nm. Results were reported as a percentage of DMSO control cells. IC50 values represent the concentration at which cell growth was inhibited by 50 %. Furthermore, for determining the time-dependent manner, A549 cells were treated with PMF (50 μM) for 12, 24, 36, 48 and 72 h, respectively; CCK–8 assay was used to determine cell proliferation inhibition.
Cell cycle analysis and apoptosis assay by flow cytometry
For cell cycle analysis, cells were cultured with either diluent control (DMSO) or various concentrations of PMF (25, 50 and 75 µM) for 48 h, and then were treated with 0.25 % trypsin to prepare the single cell suspension in phosphate buffered saline with 10 % FBS. After that, cells were washed and fixed in 70 % precooled ethanol. Finally, cells were stained by propidium iodide (PI)/RNase A and measured by flow cytometry (FACS Calibur, BD Biosciences).
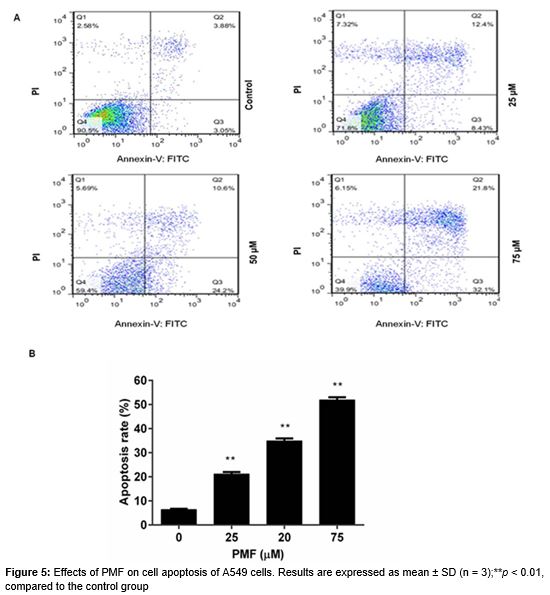
Annexin V - FITC Apoptosis Detection Kit (Beyotime, Jiangsu, China) was used to determine the effect of PMF on A549 cells apoptosis. After treatment with or without various concentrations of PMF (25, 50 and 75 µM) for 48 h, cells were harvested and then were stained by annexin-V-fluorescein isothiocyanate (FITC) and PI according to the manufacturer’s protocol. Subsequently, the apoptotic cells were measured on a FACS Calibur Flow Cytometer (BD Biosciences, San Jose, CA, USA). The cells apoptosis in early was only labeled by annexin - V while the cells apoptosis in late was labeled by both annexin-V and PI.
In vitro invasion and migration assay
Cell invasion abilities were assessed by transwell well culture chambers (Corning Incorporated Tewksbury, MA, USA) with matrigel (BD Biosciences, Shanghai, China). In brief, the lower chamber was filled with 0.75 mL of RPMI 1640 with 10 % FBS. Then 0.5 mL of A549 cells with serum-free RPMI 1640 (1 × 105 cells/well) were seeded in the upper chamber and treated with or without indicated PMF (10, 20 and 40 µM). After incubation for 48 h at 37 °C in 5 % CO2 atmosphere, the invaded cell was stained with hematoxylin-eosin solution and counted by the optical microscope (Olympus, Japan).
For transwell migration assay, the lower chambers were filed with RPMI 1640 medium with 10 % FBS and 5 × 104 cells with serum-free RPMI 1640 medium were seeded in the upper chamber with a non-coated transwell insert. And the subsequent steps were similar with the invasion experiment above. The migrated cells in the lower chamber were also stained with hematoxylin-eosin solution and observed by the optical microscope (Olympus, Japan).
Western blotting
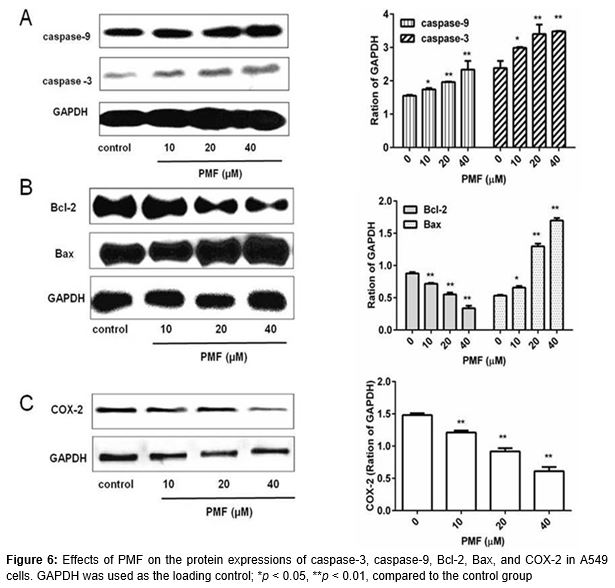
Cells were harvested after treatment with various concentrations of PMF (10, 20 and 40 µM) for 48 h. Subsequently, cells were lysed by western blot & IP cell lysis buffer (Beyotime, Jiangsu, China) and then the total protein was quantified by the BCA Protein Assay Reagent (Sangon Biotech, Shanghai, China).Equal amounts of protein (20 µg) was loaded to the sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose filter membrane. Then, the membrane was incubated with anti-caspase-3 (1: 200), anti-caspase-9 (1: 200), anti-Bcl-2 (1: 200), anti-Bax (1: 200), anti-COX-2 (1: 200) and anti-GAPDH (1: 200) antibodies at 4 °C for 12 h followed by corresponding horseradish peroxidase-conjugated secondary antibodies. Finally the immunoreactive band was visualized with ECL-detecting reagents. The primary antibodies included rabbit caspase-3polyclonal antibody, rabbit caspase-9 polyclonal antibody, rabbit Bcl-2 polyclonal antibody, rabbit Bax polyclonal antibody, rabbit COX-2polyclonal antibody, and rabbit GAPDH polyclonal antibody, which were purchased from Santa Cruz Biotechnologies (Santa Cruz, CA).The secondary antibody was horseradish peroxidase-conjugated goat anti-rabbit IgG and was purchased from Wuhan Boster Biological Technology (Wuhan, China).
Statistical analysis
All Data was expressed as mean ± standard deviation of three independent experiments and p < 0.05 was considered statistically significant. Data analysis was carried out using SPSS 19.0 software package and one way analysis of variance (ANOVA) with Dunnett’s test was used to compare the means between two groups.
Results
PMF inhibits proliferation of A549 cell in vitro
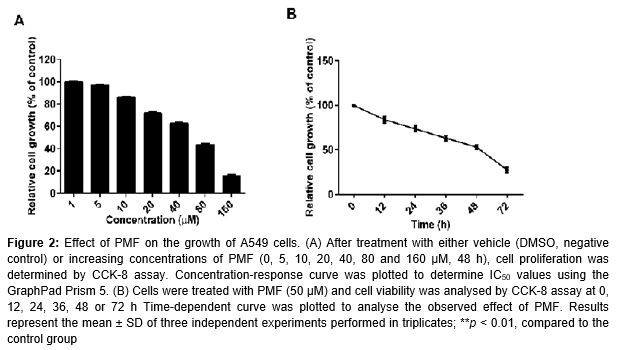
The effect of PMF on A549 cell viability was detected by the CCK-8 assay. As shown in A, the estimated IC50 of PMF was 54.51 µM in A549 cells. Compared with the vehicle group (DMSO group), PMF induced significant growth inhibition in concentration-dependent and time-dependent manners in 48 h (B).
PMF inhibits invasion and migration of A549 cell in vitro
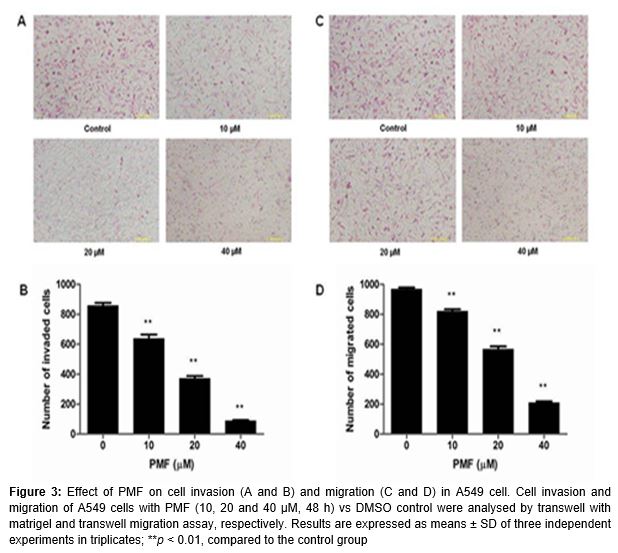
We confirmed the inhibitory effect of PMF on A549 cells invasion and migration by the matrigel invasion assay and the transwell migration assay in vitro, respectively. A and B showed the effect of PMF on A549 cells invasion and there is a significant difference between the control and PMF treated groups (p < 0.01). Furthermore, as shown in C and D, PMF at 10, 20 and 40 µM also effectively inhibited lung cancer cell migration (p < 0.01).
PMF induces cell cycle arrest and apoptosis in A549 cells
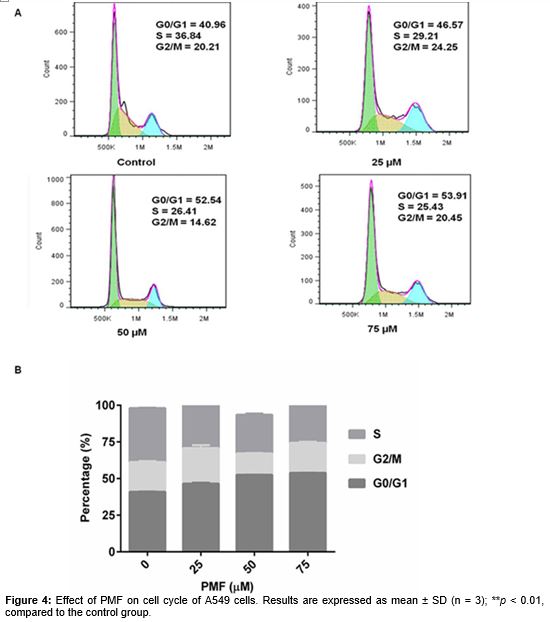
To investigate whether the anti-proliferative effect of PMF is due to cell cycle arrest and/or apoptosis, we measured the effect of PMF on cell cycle progression and apoptotic response in A549 cell lines. Cells were treated with either increasing concentrations of PMF (25, 50 and 75 µM) or vehicle control (DMSO) for 48 h and cell cycle distributions were determined by PI staining and flow cytometry. Upon incubation with PMF, A549 cells showed a dose-dependent accumulation in G0/G1, indicating cell cycle arrest in this phase (A and B).
Apoptotic response was determined by both annexin V and PI staining and flow cytometry in A549 cells. After 48 h exposure to PMF, the cell apoptosis rate was increased from 21.16 % to 51.93 % with increasing concentrations of PMF (25, 50 and 75 µM), compared with the control group (A and B).
Protein expression levels of caspase-3, caspase-9, Bcl-2, Bax and COX-2
Based on the results above, PMF inhibited the proliferation, migration and invasion of A549 cells and induced cell cycle arrest and apoptosis. To further explore the possible mechanisms, related proteins were detected by western blotting. As shown in , it is interesting that the protein expression levels of caspase-3, caspase-9, and Bax were significantly up-regulated while Bacl-2 and COX-2 expressions were down-regulated. All these results indicated that the anti-proliferation effect of PMF on the lung cancer cell might be related to promoting apoptosis proteins expression.
Discussion
Apoptosis is a mode of animal cell death which is found in normal tissues and tumors. Apoptosis is essential for normal cell development and disturbance of apoptosis is one of the important reasons the development of tumors [12]. In our present study, we found that PMF could significantly inhibit the proliferation of human non-small cell lung cancer (A549 cell) and induce cell apoptosis, which suggested that the anti-proliferation effect might be related to the enhancement of apoptosis.
Based on previous researching data, two alternative but overlapping pathways (the extrinsic and intrinsic pathways) cause cell apoptosis [13]. The extrinsic pathway can be activated by death receptors while the intrinsic pathway can be activated by the release of mitochondrial proteins [14,15]. Caspase-8 has been demonstrated to play a crucial role in the activation of the extrinsic pathway and activated caspase-9 is essential for activating the intrinsic pathway [13]. Both activated caspase-8 and caspase-9 can activate executioner caspases like caspase-3 which will activate death substrates and lead to cell death [16]. Caspase-3 is a key executor in the process of apoptosis and can hydrolyze specific protein substrates [17]. Caspase-3 and caspase-9 expressions are often suppressed in various human cancers such as lung cancer, breast cancer, prostate cancer [18-20].
In addition, the Bcl-2 protein family is an important regulator of the release of pro-apoptotic mitochondrial proteins. Bcl-2 is a crucial anti-apoptotic protein and Bcl-2 over-expression contributes to cell survival. The up-regulation of Bcl-2 expression is found in non-small-cell lung cancer [22]. Furthermore, Bax is a pro-apoptotic member of Bcl-2 family and might act on the mitochondrial membrane [22]. Importantly, our result showed that PMF could enhance caspase-3, caspase-9 and Bax expressions while it decreases Bcl-2 expression. Thus, we found that PMF induced lung cancer cells apoptosis which might be related with activation of mitochondria-mediated apoptosis pathway.
Furthermore, cyclooxygenase 2 (COX-2) is a key enzyme in aberrant arachidonic acid metabolism and can catalyze the formation of prostaglandin E2 (PGE2) [23]. It has been reported that COX-2/ PGE2 can promote lung cancer invasion/migration and high expression of COX-2 is often found in lung cancer [24]. In addition, previous data showed that PGE2 activate Bcl-2 and interleukin-6 (IL-6) which can inhibit cell apoptosis and increase cancer cell invasion and migration, respectively [25]. In our present study, PMF significantly decreased cell invasion and migration, and reduced the protein expression level of COX-2 in A549 cells, which indicated that the inhibitory effect of PMF on lung cancer invasion and migration might be associated with decreased COX-2 expression in vitro.
Conclusion
The findings of this study show that PMF inhibits the proliferation, migration and invasion of human non-small cell lung cancer A549 cells, and also induces cell apoptosis. The mechanism may be related to the activation of the intrinsic mitochondrion-mediated apoptosis pathway by down-regulating the expression of Bcl-2 and up-regulating the expression of caspase-3, casepase-9 and Bax. Furthermore, PMF probably influences COX-2 expression and inhibits cell growth.
References
Archives


News Updates