S C Ooi1,
C C Ho2,
H F Seow1 ![]()
For correspondence:- H Seow Email: shf@putra.upm.edu.my Tel:+60389472387
Received: 4 July 2015 Accepted: 13 April 2016 Published: 31 July 2016
Citation: Ooi SC, Ho CC, Seow HF. Isolation of a potential anticancer agent with protein phosphatase inhibitory activity from soil-derived Penicillium sp strain H9318. Trop J Pharm Res 2016; 15(7):1423-1429 doi: 10.4314/tjpr.v15i7.10
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To determine the effect of the secondary metabolites from Penicillium sp. H9318 on cytotoxicity and cell cycle progression.
Methods: A yeast PP1 inhibitory screening system was carried out to confirm the presence of anti-PP1c activity in crude acetone extracts of strain H9318. The extracts were fractionated and identified as Fraction S1 and Citrinin 9318 (CTN9318). Various cancer cell lines were used to test for the toxicity of the crude acetone extracts, Fraction S1 and Citrinin 9318, using MTT viability assay.
Results: It was found that a colorectal cancer cell line, HT-29, was susceptible to Fraction S1 and Citrinin 9318. A propidium iodide (PI)-incorporated DNA assay was used to show that there was G2/M arrest in HT-29 by Citrinin 9318.
Conclusion: Citrinin 9318 inhibits the viability of HT-29 via mitotic block. The results suggest that Citrinin 9318 is capable of exerting cytotoxicity and mitotic arrest in a colon cancer cell line, HT29.
Introduction
Microfungi H9318 has been identified as a Penicillium sp. isolated from the soil under leaflets at Maliau Basin, Sabah reserved forest. Preliminary experiments showed that ethyl acetate extracts from H9318 strain contain a PP1-inhibitory component (Fraction S1) that is useful for the study of the mode of action of H9318 strain when exposed to cancer cell lines.
PP1 is one of the serine/threonine protein phosphatases belonging to the PPP gene family which is abundantly expressed in animal cells and is able to manipulate cell cycle and apoptosis. Various natural inhibitors for PP1, okadaic acid and calyculin A, have been discovered to disrupt cell cycle and progressed into apoptosis in an oncogenic transformed and epithelial tumour lines [1,2].
Citrinin (CTN) is a mycotoxin found in food and feed as a contaminant and it can cause nephrotoxicity and genotoxic to animal and yeast cells [3,4]. Other studies have shown that CTN is also capable to induce cell cycle arrest in a mouse skin [5] and apoptosis in HL-60, a human promyelocytic leukemia cell line. CTN9318 was isolated from H9318 strain but in contrast to Fraction S1, it did not yield anti-PP1c effect. CTN9318 has a similar structure with the commercially available Citrinin. The objectives of our studies were to find out whether anti-PP1 effect of Fraction S1 could trigger cell cycle arrest and the potential genotoxic effect of CTN9318 on the induction of cell cycle arrest on colon cancer cell line, HT-29.
Methods
Microfungi culture and preservation
Microfungi H9318 was cultured in a Potato dextrose agar (PDA). PDA medium was prepared by adding 2 g of the PDA powder (Merck) into 50 ml of distilled water and autoclaved at 121 oC for 20 min. Agar medium (25 mL) was poured into a Petri dish with 25 mL and left to harden before inoculation. Conidia of the H9318 strain were streaked onto the surface of the agar using a sterile inoculating wire loop and incubated at 25 oC to yield single colony. After 5 days of incubation, greenish and powdery appearance of the cultures indicated mature growth of the microfungi. Cultures were sub-passaged onto a slant agar and kept at 4 oC for future experimental use. Conidia were transferred to a Bijou bottle containing sterile water kept at 4 oC for short term storage and also transferred to a slant agar kept at -20 oC for long term storage.
Fermentation culture of H9318 strain
Fermentation of H9318 was done in the laboratory with the YPD liquid medium to yield secondary metabolites. The YPD liquid medium consists of yeast extracts (Sigma), Peptone enzymatic digested from meat (Fluka), Sucrose (Amresco), KH2PO4 (Fluka) and MgSO4.7H2O (Amresco) with a pH of 5.5. The medium (25 mL) was poured into 100 mL conical flasks and cotton-sealed prior to autoclaving. Then, conidia of H9318 were inoculated into each conical flask and placed in a refrigerated shaking incubator (IS-971R, Lab Companion, Jeio Tech Inc.) at 28 oC, 220 rpm for 120 h.
At day 5 (120 h), cultures were filtered through a filter paper (Whatman No.1) by using a glass funnel and collected into 1 L Erlenmeyer flask. Acetone was added into the culture medium at 1:1 ratio. Acetone breaks cell wall and plasma membrane besides extracting bioactive secondary metabolites. The mixture was gently swirled and kept at 4 oC for 3 days to ensure a complete extraction. Next, acetone was evaporated from the mixture using Eyela rotary evaporator N-1000, Eyela oil bath OSB-2000, Eyela pump A-1000S, at 42 oC. The remaining liquid subjected to freeze drying to collect the crude freeze-dried acetone extract.
Equal volume of ethyl acetate was poured into the crude acetone extract, gently swirled and kept at 4 oC for 3 days. This step was called liquid-liquid extraction (LLE) that is to separate particles into the organic or the water layer extracts. After 3 days, two visible immiscible layers were observed and the ethyl acetate layer was separated out by a glass separator and the ethyl acetate layer extract was evaporated using a rotary evaporator at 42 oC. The process of evaporation ceased when brownish extracts were observed on the collecting flask.
HPLC collection and fractionation of Fraction S1
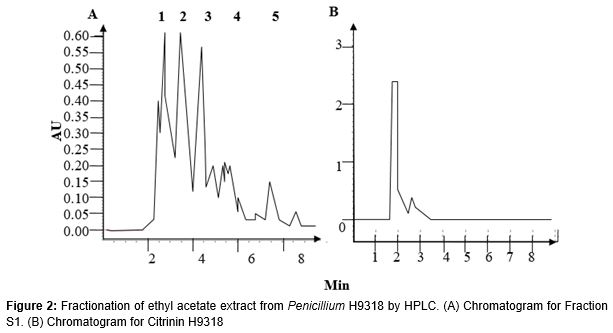
The purified ethyl acetate extracts were dissolved in 100 % methanol and filtered through a microporous membrane filter (Acrodisc LC, 0.20 µm, 13 mm, GHP, PALL) into an amber screw neck vial 12 x 32 mm with bonded PTFE/Silicone membrane. The vial was put into the HPLC machine and the sample analytes (50 µL) were injected into the machine.
The HPLC machine consists of Waters 2695 Separation Module, Waters 2487 UV detector channel and Waters Fraction Collector II. A semi-preparative reverse phase column (Licospher C18, Merck) was used with a flow rate of 5 ml/min. A running time of 15 – 30 min was set with an isocratic mode using acetonitrile and water which were programmed at the ratio of 40:60. Millennium 32 software was used.
Freeze-drying process
Collected fractions from HPLC were pooled in Eppendorf tubes and distilled water added in order to allow the fractions to be frozen. The Eppendorf tubes were put into the freeze-drying flasks and stored at -80 oC overnight before placing in the freeze-dryer (Telstar).
Freeze-drying process was performed in a lyophilization equipment which consists of a drying chamber with temperature controlled shelves, a condenser to trap water removed from the product, a cooling system to supply refrigerant to the shelves and condenser, and a vacuum system to reduce pressure in the chamber and condenser to facilitate the drying process. The freezing condition was a prerequisite from the solid phase to gas phase at low pressure. Freeze-dried fractions were kept at -20 oC to avoid degradation.
Preparation of treatments
Crude freeze-dried acetone extract was dissolved in RPMI medium. Fraction S1 from HPLC elution at 4 min as in Ong et al [6] was dissolved in less than 0.5 % of DMSO and diluted with RPMI 1640 medium upon cell treatment. CTN9318 was dissolved in < 0.5 % of ethanol and DMSO, was then diluted with RPMI 1640 medium. All the samples were filtered through microporous membrane filters (PES membrane, 0.22 µm, Millipore) and kept at -20 oC.
Yeast PP1 inhibitory screening system
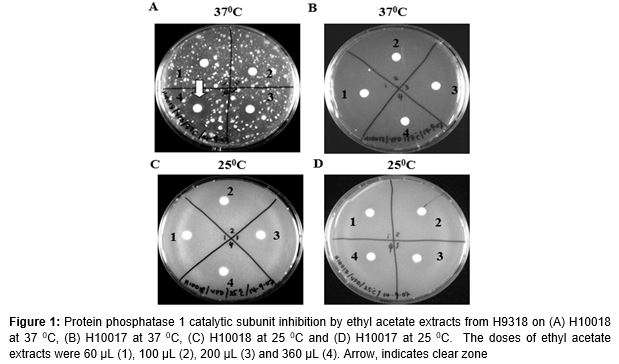
Yeast testers, Saccharomyces cerevisiae – H10018, a wild-type yeast carrying a GLC gene encoding for PP1c, or was designated as PAY704-1 and H10017, a mutant yeast carrying a high temperature sensitive allele, glc-10, or was designated as PAY700-4. Were used for screening both types of yeasts were grown in a Yeast Peptone Dextrose (YPD) medium. Yeasts were cultured in a sterile liquid medium (YPD) at 25 oC in a refrigerated shaking incubator and agitated at 220 rpm. After 3 days of incubation, 100 µL of the yeasts were added into a 25 mL of YPD agar and were poured into a Petri dish. After that, a 6 mm paper disc was soaked with the treatment fraction and was placed onto the surface of YPD agar. After all the paper discs were arranged on the agar, the Petri dishes were incubated at 25 and 37 oC. After 3 days, inhibition zones were measured.
Cell culture
Breast (MCF-7 and MDA-MB-231), nasopharyngeal (CNE1, TWO1 and TWO4), cervical (Hela), ovarian (CaOV3), prostate (PC3) and colorectal cancer (HT-29) cell lines were obtained from ATCC. Cells were grown in RPMI 1640 medium (Gibco) supplemented with 10 % FBS and 1 % penicillin/streptomycin (Gibco). Cells were cultured and propagated at 37 oC in a CO2 incubator with 5 % CO2 in a humidified atmosphere.
Cell count after trypsinization for bioassays
HT-29 cells were trypsinized with 0.25 % of Trypsin-EDTA (Gibco) after 80 % confluency. Then, the suspension of cells was diluted with Trypan blue dye and counted using a haemocytometer. Cells were visualized and counted under a phase-contrast microscope.
MTT viability assay
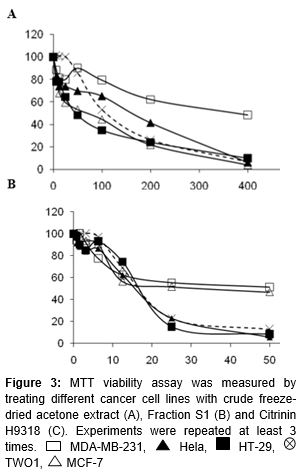
Seven cell lines were harvested and treated with two-fold serial dilutions of Fraction S1 (400 µg/mL) and CTN9318 (50 µg/mL) in three separate experiments as shown in . Each triplicate of the treatments was compared to the corresponding untreated controls. Next, 10 µL of MTT solution (5 mg/mL) was added into each well using a multichannel pipette and incubated for 4 h in a CO2 incubator. Then, the liquid in each well was discarded and the formazan crystals were solubilized with 100 µL of DMSO. The microplate was mixed properly and absorbance was measured using an ELISA reader (DYNEX Technologies MRX11) at 570 nm wavelength. Experiments were repeated at least three times and data were expressed as percent cell viability against the concentrations (µg/mL).
PI-incorporated DNA assay
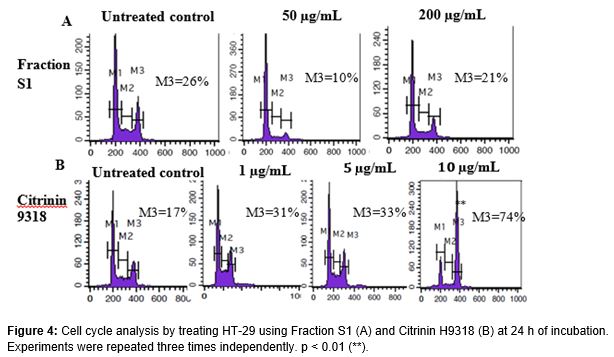
Mitotic arrest was determined by using the Propidium iodide (PI) staining intercalated with the DNA content in a cell and analyzed by a flow cytometer, FACSCalibur (Becton Dickinson). HT-29 cells with a concentration of 1.5 x 105 cells/mL were seeded into a 24-well plate and incubated overnight. Cells were exposed to 50 and 200 µg/mL of Fraction S1 and 1, 5 and 10 µg/mL of CTN9318. After 24 h of incubation, the cells were harvested and washed with PBS/0.3 % of BSA and were pelleted at 2000 rpm, 5 min. Later, the pellets were fixed with ice-cold 70 % of ethanol and kept overnight. The next day, the fixed cells were washed and stained with PI after adding RNase solution. Percent of the PI-stained cells (10,000 gated single cells) were calculated using CELLQuest Pro software.
Statistical analysis
Statistical analysis was performed using Student’s t-test (SPSS version 12.0) to evaluate differences between treated and untreated groups. Statistical significance was defined as p < 0.05 (*) and p < 0.01 (**).
Results
Yeast PP1 inhibition
The viability of two Saccharomyces cerevisiae strains, H10017 and H10018 were tested using ethyl acetate extracts from strain H9318. This assay system was carried out to determine whether the extracts from strain H9318 could inhibit PP1c.
H10017, also known as PAY700-4, is a mutant yeast. It contains a temperature sensitive allele glc7-10 which cannot survive at the restrictive temperature of 37 oC. H10018, also known as PAY704-1, is a wild-type yeast that carries a normal GLC gene encoding PP1c. Our results show that ethyl acetate extract from H9318 strain killed the wild type yeast, H10018 at 37 oC in a dose-dependent manner (A). The higher the amount of the extract, the higher the number of H10018 killed. There was a clear inhibition zone indicated with arrow. As compared to Petri dish C, there was no inhibition zone observed at 25 oC, showing that the extract inhibited PP1c specifically. This result is compatible with the other researchers in the Yeast PP1 screening assay which sets a key point for our laboratory to reproduce the extracts for our next bioassay experiments [6].
Fraction S1 and CTN9318
The ethyl acetate extract was dissolved in 100 % methanol and 10 µL was injected into the semi-preparative column. The running time was 15 min in order to have a maximum display of the peaks separated by the HPLC column. There were five prominent peaks observed in the chromatogram (1, 2, 3, 4 and 5 as indicated in A) between the running time from 0 to 10 min. The highest peak detected at 4 – 4.5 min was designated as Fraction S1 (indicated as 2 in the chromatogram). No contaminant occurred as methanol did not contain substance that would be detected or separated by HPLC. The chromatogram was compatible with Ong et al as Fraction S1 was eluted at 4 min [6]. B shows a single and prominent peak of CTN9318 compared at different wavelengths. It was ascertained as a pure compound isolated from H9318 strain which possessed a similar structure to Citrinin available in the market. Structural determination was done by Dr. Chang’s student. CTN9318 is a gift from Dr. Leng Chee Chang of Hawaii University.
Cell viability
Cell viability of various cancer cell lines was calculated using crystal violet formazan dye as detected at 570 nm by an ELISA reader. The MTT assay shows that Fraction S1 inhibited HT-29 with an IC50 ~ 50 µg/mL (A). IC50 for MCF-7, TWO1 and Hela were 80 µg/mL, 110 µg/mL and 180 µg/mL in response to Fraction S1. This is the first report for the cytotoxic results of Fraction S1 on cancer cell lines. CTN9318 exerted a lower IC50 with ~ 18 µg/mL on HT-29 (3-fold lower than Fraction S1) if compared to Fraction S1. The cytotoxic effect of CTN9318 on cancer cell lines could also be found on the report of the other laboratory [7] which has highlighted the IC50 of CTN9318-treated MCF-7 at 18 µg/mL similar with our cytotoxic data. The highest concentration used in CTN9318 was 8-fold lower than that for Fraction S1, showing that CTN9318 was more potent than Fraction S1 (B).
PI-incorporated DNA
shows that PI staining was detected by a FL-2 detector in BD FACSCalibur. The DNA content was calculated as % for the gated HT-29 cells. CTN9318 arrested HT-29 at G2/M phase after 24 h of incubation at 1, 5 and 10 µg/ml (B). It shows a dose-dependent increment of the PI-stained DNA cells at G2/M phase. For other studies, Citrinin (12.5 µg/ml) could trigger a 36.7 % increase of sub-G1 HL-60 cells together with our PI-incorporated DNA assay [8], we determined that CTN9318 was genotoxic to cancer cell line HT29 and HL60. Fraction S1 did not exert cell cycle arrest as compared to CTN9318 (A).
Discussion
Yeast PP1 inhibitory screening system has shown the presence of PP1c inhibitors from the extracts of strain H9318. The ethyl acetate extracts of H9318 strain contain anti-PP1c property. PP1c is important for the yeasts to survive in YPD media as it is utilized by both the mutant H10017 and wild-type H10018 to mediate a signaling pathway for cell-wall integrity which is essential for yeast survival [9]. However, the glc7-10 allele of H10017 does not permit survival at the high temperature of 37 oC as shown by B because the cell wall integrity is disrupted at 37 oC.
To ascertain specificity, a control set at 25 oC is needed. It is of utmost importance that ethyl acetate extracts from strain H9318 do not yield positive results (shown by the appearance of inhibition zones) on yeast H10018 at 25 oC. This is due to the fact that at 25 oC, there are more than one cell wall integrity pathways such as the bck1 and Mpk1 signalling pathways that enable survival of yeasts on agar besides the PP1c-regulated pathway. The appearance of inhibition zone at 25 oC means that the screening result is negative for PP1c inhibitors as only the disrupters of PP1c signaling pathway are selected for the next phase of the study.
After the preliminary screening for anti-PP1c, the biological effect of crude acetone extracts on various cancer cell lines was investigated. The MTT assay results had shown that HT-29 cells were inhibited by both Fraction S1 and CTN9318 in a dose-dependent manner. The morphological appearance of HT-29 cells treated with CTN9318 was round, bright and still adherent while it was irregular in shape and dull when treated with Fraction S1 (data not shown). Thus, this indicated that the effect of Fraction S1 and CTN9318 from H9318 strain was different.
For the cell cycle analysis, HT-29 treated with CTN9318 was arrested at G2/M phase and CTN9318 was genotoxic to HT-29 cells. Further evaluation needs to be carried out to determine which cell cycle proteins were affected by the treatment of CTN9318 and the stark difference in the activity of Citrinin on the cell cycle progression of HL60 and HT-29 shows that CTN9318 might contribute to premature mitosis of the adhered HT-29 cells rather than the suspended HL60 cells. Fraction S1 failed to trigger a G2/M arrest in HT-29 showing that PP1 inhibition is not necessary for the mitotic disruption in the event for cell death. Further studies on involvement of the other proteins in the mitotic arrest induced by CTN9318 and mechanism of cell death induced by Fraction S1 needs to be carried out.
Conclusion
The findings of this study indicate that ethyl acetate extract of H9318 strain Penicillium contains a PP1c inhibitor, Fraction S1. The results also show that the genotoxicity of CTN9318 can arrest HT-29 at G2/M phase and the possible PP1-inhibitory cell death mechanism triggered by Fraction S1 on HT-29 does not involve cell cycle arrest at G2/M phase. In a future study, efforts will be made to combine the two extracts isolated from H9318 strain on HT-29 cell line for the further apoptotic analysis.
Declarations
Acknowledgement
References
Archives


News Updates