Ruchadaporn Kaomongkolgit1 ![]() ,
Ichaya Yiemwattana2
,
Ichaya Yiemwattana2
For correspondence:- Ruchadaporn Kaomongkolgit Email: ruchadapornk@nu.ac.th Tel:+665966065
Received: 19 June 2016 Accepted: 18 September 2016 Published: 31 October 2016
Citation:
Kaomongkolgit R, Yiemwattana I.
α-Mangostin suppresses 12-o-tetradecanoylphorbol-13-acetate-mediated matrix metalloproteinase-9 ex
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the effect of α-mangostin on matrix metalloproteinase (MMP)-2 and MMP-9 ex
Methods: Cytotoxicity of α-mangostin on U2OS cells was determined by MTT assay. MMP-2 and MMP-9 activities, and mRNA ex
Results: U2OS viability was significantly decreased when cells were exposed to α-mangostin at 2.5 and 5 µg/mL compared to the untreated control (p < 0.05). α-Mangostin inhibited MMP-2 and MMP-9 activities stimulated by TPA at concentrations of 1.0, 1.5, and 2.0 µg/mL. MMP-9 mRNA ex
Conclusion: This is the first study reporting the inhibitory effects of α-mangostin on TPA-mediated MMP-9 ex
Introduction
Osteosarcoma is the most common primary bone malignancy found in children and young adults. It usually occurs in the distal long bones, but can also affect other bones in the body [1]. Chemotherapy is one of the most essential treatments for osteosarcoma. Although a number of chemotherapeutic agents have been used to treat osteosarcoma, survival rate of patients has not been substantially improved [2]. One of the primary reasons for the failure of current treatments is that patients with stage IV cancer often develop resistance to anticancer agents [3]. Matrix metalloproteinases (MMPs) are a major group of enzymes that regulate cell-matrix composition. Gelatinases (MMP-2 and MMP-9) are members of MMP family that plays an important role in the progression and metastasis of human cancers [4]. Previous reports have showed that MMP-2 and MMP-9 were identified in osteosarcoma tissue and can participate in the progression of osteosarcoma [5]. In addition, a recent study demonstrated that MMP-2 overexpression was related to a poor prognosis [6] and high MMP-9 expression were significantly associated with lower overall survival compared to their counterparts with low or undetectable MMP-9 expression [7].
α-Mangostin is a plant-derived antitumor xanthone isolated from Garcinia mangostana Linn. or mangosteen. This agent possesses a variety of pharmacological activities including anticancer, cytotoxicity, antioxidant, anti-inflammation, antihistamine, antibacterial activity, antifungal activity, anti-HIV, antiulcer, and CNS depressant activities [8]. In vitro and in vivo studies demonstrated that α-mangostin displayed a broad antitumor activity against various human cancer cells [9]. In addition, α-mangostin was able to induce apoptotic cell death in canine osteosarcoma D-17 cells [10] and inhibited metastasis of human cancer cells [11-16]. However, there was no data about the effect of α-mangostin on the MMP expression in human osteosarcoma cells.
The purpose of this study was to investigate the effect of α-mangostin on MMP-2 and MMP-9 expression in U2OS human osteosarcoma cell lines in vitro.
Methods
Preparation of α-mangostin
α-Mangostin was received from Department of Pharmacognosy and Pharmaceutical Botany, Faculty of Pharmaceutical Sciences, Chulalongkorn University, Thailand. The purified α-mangostin was prepared according to the protocol of a previous study [17]. The pericarp of mangosteen was macerated in hexane to remove nonpolar compounds and was subsequently macerated in ethyl acetate. α-Mangostin was obtained from the crude extract by chromatography on a silica gel column and elution with hexane/ethyl acetate (4:1). The selected fraction was identified as α-mangostin using mass spectrometry, nuclear magnetic resonance spectroscopy, and a Gallenkamp melting-point apparatus.
Cell culture
Human osteosarcoma cell line (U2OS) was kindly provided by Professor Dr. Prasit Pavasant (Faculty of Dentistry, Chulalongkorn University). Cells were cultured in Dulbecco’s modified Eagle’s medium supplemented with 10 % fetal bovine serum (FBS), 2 mM L-glutamine, 100 IU/mL penicillin, 100 µg/mL streptomycin and 5 µg/mL amphotericin B at 37 ºC in humidified atmosphere of 95 % air, 5 % CO2. Medium and supplements were purchased from Gibco® (Grand Island, NY, USA).
MTT assay
Cytotoxic effect of α-mangostin and 12-O-tetradecanoylphorbol-13-acetate (TPA) (Sigma®, St. Louis, MO, USA) on U2OS cells were determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) test (Sigma®, St. Louis, MO, USA). MTT assay is based on the reduction of the yellow tetrazolium salts to purple formazan crystals by dehydrogenase enzymes secreted from the mitochondria of metabolically active cells. The amount of purple formazan crystal is proportional to the number of viable cells. MTT assay was performed according to the manufacturer’s instruction. Cells were seeded at 5 × 104 cells per well in 24-well plate. The cells were serum-starved and then treated with various doses of α-mangostin (0, 0.5, 1.0, 1.5, 2.0, 2.5 and 5 µg/mL) in serum free culture medium as previously mentioned. After 24 or 48 h, the culture medium was aspirated, replaced with 0.5 mg/mL MTT solution and incubated for 30 min at 37 ºC in a 5 % CO2 humidified atmosphere. The solution was then aspirated and 1000 µL of DMSO (Sigma®, St. Louis, MO, USA) was added to dissolve the formazan crystals. The absorbance was measured at 570 nm using Genesis10 UV-vis spectrophotometer (Thermo Spectronic, NY, USA). Viable cell number was calculated from the standard curve of cell number [15].
Gelatin zymography
Gelatin zymography was used to determine MMP-2 and MMP-9 activities secreted from U2OS cells. Briefly, conditioned media were collected and resolved in 10 % sodium dodecyl sulfate polyacrylamine gel electrophoresis (SDS-PAGE) gels copolymerized with 1 % gelatin. Then, Gels were washed three times with 2.5 % Triton X-100 and incubated in buffer containing 50 mM Tris, 10 mM CaCl2, 0.15 M NaCl, and 0.02 % Briji-35 at 37 ºC overnight. Finally, Gels were stained with 0.1 % Coomassie Brilliant Blue R250 in 7.5 % acetic acid and 12.5 % methanol for 30 min, and then destained in 7.5 % acetic acid and 5 % methanol to visualize the proteolytic bands. Bands were quantified using Scion Image software (Scion Corporation, Frederick, MD, USA) [15].
Real-time polymerase chain reaction (RT-PCR)
Total RNA was isolated by Nucleospin RNAII (Macherey-Nagel GmbH & Co. KG) following the product protocol. The mRNA concentrations of the samples were determined using the Nanodrop (ND-2000c; Thermo Fisher Scientific, Wilmington, USA). cDNA was synthesized from 1 µg of total mRNA and mixed with LightCycler® 480 DNA SYBR Green I Master (Roche Diagnostics GmbH, Mannheim, Germany) and gene-specific primers (Takara Bio, Shiga, Japan) according to the manufacturer’s instructions. The PCR primer sequences of the MMP-2, MMP-9, and GAPDH were used as follows: sense primer for MMP-2, 5’-GGCCCT GTCACTCCTGAGAT-3’ and anti-sense 5’-GGCATCCAGGTTATCGGG GA-3’; sense primer for MMP-9, 5’-GACTCGG TCTTTGAGGAGCC-3’ and antisense 5’-GAAC TCACGCGCCAGTAGAA-3’, and sense primer for GAPDH, 5’-TGAAGGTCGGAGTCAACGG AT-3’ and antisense 5’-TCACACCCAT GACGAACATGG-3’. All PCR reactions were performed in duplicate using the LightCycler® 480 II Real-Time PCR System (Roche Diagnostics Ltd, Rotkreuz, Switzerland).
The cycling conditions were as follows: initial denaturation at 95 °C for 3 min, followed by 40 cycles at 95 °C for 10 s, 53 °C for 45 s. A melt curve assay was then performed (55 °C for 1 min and then the temperature was increased by 0.5 °C every 10 s) to detect the formation of primer-derived trimers and dimers. To ensure the fidelity of mRNA extraction and reverse transcription, all samples were subjected to PCR amplification with oligonucleotide primers specific for the constitutively expressed gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and normalized. Data were analyzed with the LightCycler® 480 software version 1.5. Quantification was calculated using the starting quantity of the cDNA of interest relative to that of GAPDH cDNA in the same sample [18].
Wound healing assay
To determine cell motility, U2OS cells were seeded as previously described and grown until they were completely confluent. Cells were wounded by scratching with a sterile yellow micropipette tip and washed with PBS three times. Cells in each well were then incubated with TPA (10 nM) with or without the different concentrations of α-mangostin for 24 h. Photographs were taken (Olympus DP72, Hamburg, Germany) before and after 24 h of treatment and cell migration was measured using Imaging software (CellSens, Olympus, Hamburg, Germany) [19].
Statistical analysis
All data are expressed as the mean ± standard deviation (SD, n = 3). The results were analyzed using Student’s t-test, and differences were considered statistically significant only if p < 0.05. All statistical analyses were performed by SPSS for Windows software (version 13.0; SPSS Inc, Chicago, IL, USA).
Results
Cell viability of U2OS cells treated with α-mangostin
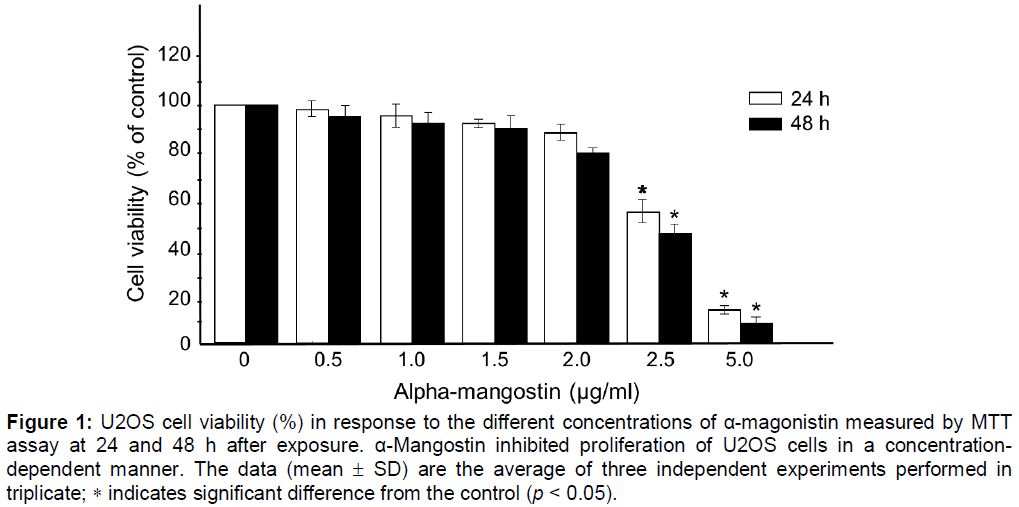
Cytotoxicity of α-mangostin was first determined by treating U2OS cells with various concentrations of α-mangostin (0, 0.5, 1.0, 1.5, 2.0, 2.5 and 5 µg/mL) for 24 and 48 h followed by MTT assay. As shown in , α-mangostin inhibited growth of U2OS cells in a concentration dependent manner. α-Mangostin at concentrations between 0 and 2.0 µg/ml was not toxic as the cell viability was not significantly altered compared to the non-treated control. However, at the concentration of 2.5 to 5 µg/mL, cell viability was significantly decreased indicating the cytotoxicity at these concentrations. As a result, α-mangostin at the concentrations ≤ 2.0 µg/mL were applied on all subsequent experiments.
Inhibition of the TPA-induced expressions of MMP-9 by α-mangostin
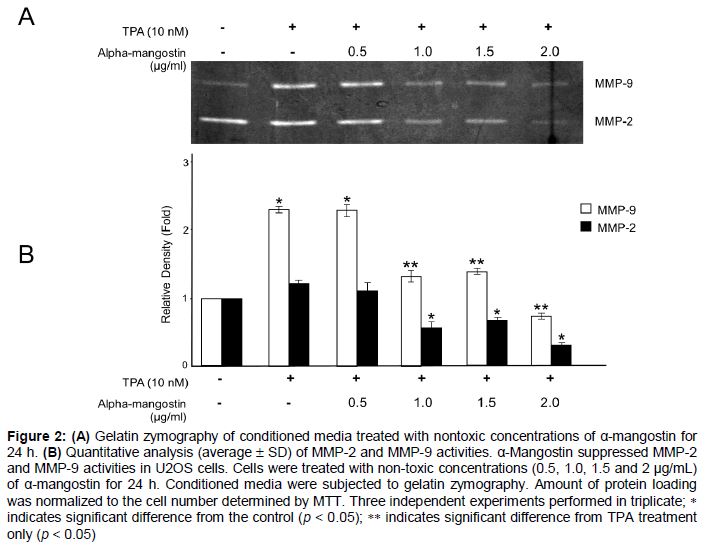
Gelatin zymograhy showed proteolytic bands of MMP-2 and MMP-9 (). Compared to the non-TPA treated cells, TPA significantly increased the MMP-9 activity. However, this effect was not observed for MMP-2. α-Mangostin significantly inhibited MMP-2 and MMP-9 activities stimulated by TPA at concentrations of 1.0, 1.5, and 2.0 µg/mL. ().
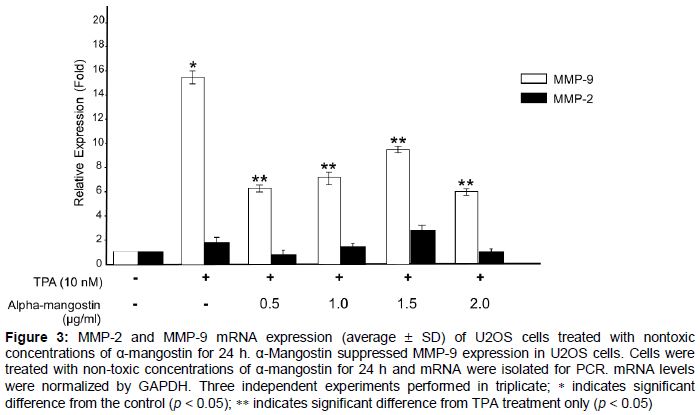
To determine whether the inhibitory effect of α-mangostin on TPA-induced MMP activity was regulated at the transcription level, real-time PCR was used. As shown in , MMP-9 mRNA expression of TPA treated U2OS was down-regulated by α-mangostin at all concentrations used. However, MMP-2 mRNA levels remained unchanged.
α-Mangostin inhibits TPA-induced migration of U2OS cells
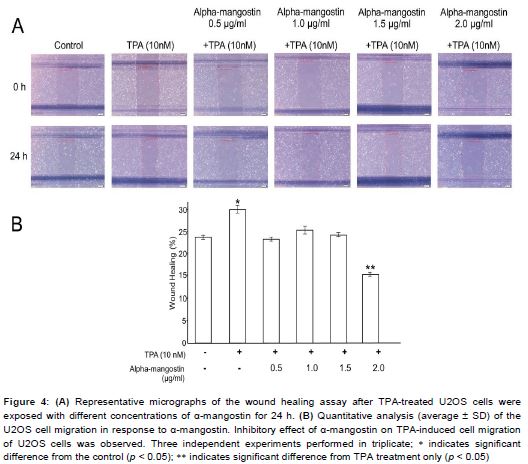
As shown in , cell motility of TPA-induced U2OS cells was significantly increased, compared to untreated cells. Treatment U2OS cells with 0.5, 1.0, 1.5 µg/mL α-mangostin reduced the motility of cells induced by TPA, while 2 µg/mL α-mangostin significantly inhibited cell motility compared to the TPA-treated U2OS.
Discussion
Despite recent advances in osteosarcoma treatment, the disease still remains a leading cause of death among cancer patients [20,21]. In addition, osteosarcoma is a highly metastatic tumor [22]. Thus, an agent that could inhibit the development of metastasis would be beneficial to patients by improving their disease-free survival time. One of important characteristics of metastasis is the migratory ability of tumor cells. Several MMPs, including MMP-2 and MMP-9 are involved in tumor migration, and increase in MMP-2 and MMP-9 protein levels and enzyme activities were observed in clinically invasive cancers [23]. Previous reports indicated that MMP-2 and MMP-9 were found in osteosarcoma tissue and they could be involved in the osteosarcoma progression [3,5].
α-Mangostin has been confirmed to consist of highly biological active molecules possessing anticancer properties such as anti-proliferative, apoptotic, anti-angiogenesis and anti-metastatic effects on human cancers [8,9,24]. Krajarng et al [10] reported that α-mangostin induced apoptotic cell death of canine OS, D-17 cells. Moreover, numerous studies indicated that inhibition of MMP expression and its activities can provide early targets for prevention of cancer metastasis [2,4]. Nevertheless, there has been no data reporting the effect of α-mangostin on the MMP-2 and MMP-9 expression in human osteosarcoma cells. To explore this effect, this study investigated the inhibitory effect of α-mangostin on TPA-induced MMP-2 and MMP-9 at the enzymatic activities and mRNA expression. Moreover, wound healing assay was also used to determine if the α-mangostin could inhibit TPA-induced migration of U2OS cells.
The results showed that α-mangostin inhibited MMP-9 activity stimulated by TPA in a dose-dependent manner. Also, α-mangostin down-regulated TPA-induced MMP-9 mRNA expression of U2OS cells. Interestingly, although α-mangostin significantly inhibited MMP-2 activity, the mRNA expression was not changed. This is possibly because α-mangostin could inhibit MMP-2 secreted from the cells (at the protein level) but it could not regulate MMP-2 expression at the transcription level
The expression of MMP-2 and MMP-9 gene is controlled by several growth factors and cytokines. By using specific inhibitors, several studies have revealed the involvement of various signaling pathways in MMP-2 and MMP-9 regulation which depends on the cell types and nature of the stimuli. Signaling pathways via MAPKs, NF-κB, and PI3K/Akt have been reported [25]. Previous studies have reported that α-mangostin showed antimetastatic effects in various human cancer cell lines through the different signaling pathways. For example, Lee et al [12] reported that α-mangostin suppresses TPA-induced MMP-2 and MMP-9 expression through the ERK signaling pathway in MCF-7 human breast adenocarcinoma cells.
Hung et al [11] reported that α-mangostin exerts inhibitory effect on metastasis of PC-3 human prostate carcinoma cells by inhibiting MMP-2, MMP-9, and urokinase-plasminogen expression through the JNK signaling pathway. Moreover, Shih et al [13] demonstrated that α-mangostin suppresses phorbol 12-myristate 13-acetate (PMA)-induced MMP-2 and MMP-9 expression via αvbeta3 integrin/FAK/ERK and NF-kappaB signaling pathway in A549 human lung adenocarcinoma cells. Therefore, investigation of signaling pathways and molecular mechanism associated with α-mangostin and MMP-2 and MMP-9 expression in osteosarcoma cells should be further investigated.
Regarding the wound healing assay, the results showed that α-mangostin significantly inhibited the TPA-induced motility of U2OS cells where compared to that of TPA treatment alone. Collectively, results from gelatin zymography, real-time PCR and wound healing assay suggest that α-mangostin prevents the transcription of MMP-9 in response to TPA. Furthermore, the anti-migration effect of α-mangostin in U2OS cells was related to the inhibition of enzymatically degradative process of tumor migration. As a consequence, α-mangostin may possibly be used for suppressing tumor migration of U2OS cells at non-cytotoxic concentrations. However, metastasis of cancer cells involves multistep processes and various cyto-physiological changes. Therefore, other methods such as cell-matrix adhesion assay, invasion assay, and in vivo model need to be further explored to understand the mechanism of inhibition on osteosarcoma metastasis.
Conclusion
This is the first study reporting the inhibitory effects of α-mangostin on TPA-mediated MMP-9 expression and TPA-induced migration of U2OS human osteosarcoma cells. The findings suggest that α-mangostin has a protective effect against TPA-mediated cell migration of U2OS via the suppression of TPA-mediated MMP-9 in vitro and thus the compound is a potential therapeutic agent for the treatment of osteosarcoma.
Declarations
Acknowledgement
References
Archives


News Updates