Hamide Hatamihanza1,
Mehrdad Hashemi1 ![]() ,
Azim Akbarzadeh2,
Fatemeh Fotouhi3,
Behrokh Farahmand3,
Hasan Ebrahimi Shahmabadi4
,
Azim Akbarzadeh2,
Fatemeh Fotouhi3,
Behrokh Farahmand3,
Hasan Ebrahimi Shahmabadi4
For correspondence:- Mehrdad Hashemi Email: mhashemi@iautmu.ac.ir Tel:+822006660
Received: 18 May 2016 Accepted: 11 October 2016 Published: 28 November 2016
Citation: Hatamihanza H, Hashemi M, Akbarzadeh A, Fotouhi F, Farahmand B, Shahmabadi HE. Generation of truncated recombinant form of tumor necrosis factor receptor-1 to produce cancer vaccine. Trop J Pharm Res 2016; 15(11):2345-2350 doi: 10.4314/tjpr.v15i11.7
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To produce truncated recombinant form of tumor necrosis factor receptor 1 (TNFR1), cysteine-rich domain 2 (CRD2) and CRD3 regions of the receptor were generated using pET28a and E. coli/BL21.
Methods: DNA coding sequence of CRD2 and CRD3 was cloned into pET28a vector and the corresponding protein was expressed under induction of isopropyl β-D-1-thiogalactopyranoside (IPTG) as 6×His tagged using E.coli BL21 (DE3) ex
Results: Protein ex
Conclusion: A truncated form of TNFR-1 has been successfully expressed in a bacterial ex
Introduction
Tumor necrosis factor-α (TNF-α) is a multi-functional cytokine with various roles in inflammation, immunity, cell proliferation, differentiation and apoptosis [1]. TNF-α exists in two forms: transmembrane and soluble which exert their effects through binding and activating two TNF family receptors: TNF receptor 1 (TNFR1) and TNF Receptor 2 (TNFR2) [2]. Many studies have been conducted on antitumor functions of the receptors [3]. The major difference between the two receptors has been reported to be the presence of a death domain in TNFR1 and absence of a death domain in TNFR2 [4]. TNF has been found to activate proapoptotic signaling cascade through TNFR1 [5] which is expressed in various types of cells, tissues [6] and tumors [7]. Studies have shown that antibodies against TNFR1 mimic the effect of TNF-α [8,9] and as a result initiate the apoptosis process. The extracellular region of the receptor consists of four cysteine-rich domains (CRDs): CRD1, CRD2, CRD3, CRD4. CRD2 and CRD3 play important roles in ligand binding [10]. TNF-α binding to TNFR1 induces receptor trimerization and recruitment of various signaling proteins. TNFR1 transduces apoptotic and anti-inflammatory signals via the employment of FAS-associated death domain protein (FADD) and further recruitment of caspase-8. Activated caspase-8 triggers a proteolytic cascade that entangles caspases-3, -6 and -7, eventually inducing apoptosis. Also TNFR-1 mediates anti-apoptotic and inflammatory responses like activation of the necrosis factor (NF)-κB by employing TNF-receptor-associated factor 2 (TRAF2) and receptor-interacting protein 1 (RIP1) [11]. Therefore, inhibition of NF-κB activity enhances TNF-α-induced cell death while an increase in NF-κB activity protects the cells from TNF-α-induced death [12].
In this study, CRD2 and CRD3 regions in a pET vector were cloned using E.coli/BL21 and expressed. Various techniques including agarose gel electrophoresis, sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), western blot, Ni-NTA chromatography, dialysis membrane, and Bradford were used.
Methods
Materials
Diaminobenzidine (DAB) was purchased from Sigma Company (USA). Acrylamide, sodium dodecyl sulfate and agarose were obtained from Merck Company (Germany). Original pET28a and recombinant pET28a plasmid were gifted and E.coli/BL21 was prepared from Pasteur Institute of Iran, Tehran. BamHI, Sal I, restriction enzymes and T4 DNA Ligase were purchased from Fermentas Company (Finland). DNA extraction kit and Ni-NTA agarose resin were obtained from Bioneer Company (South Korea) and Qiagen Company (Germany), respectively. Distilled water was used throughout the experiment. All materials were of analytical grade.
Cloning of CRD2 and CRD3
Recombinant pET28a plasmid resistance to kanamycin, containing CRD1, CRD2, CRD3 and CRD4 was transferred into susceptible bacteria (E.coli/BL21) by chloride calcium and heat shock methods. For this purpose, cultured bacteria in the log phase (OD=0.4) were centrifuged (9000 rpm, 3 min) and sediment was obtained. The sediment was mixed with 1 mL 0.1M cold calcium chloride, and placed on ice for 30 min and then centrifuged. The supernatant was then discarded and the process was repeated. The suspension of bacteria was then incubated with recombinant vector (1.5 µl, 60 ng/µl) on ice for 30 min. The mixture was shocked with temperature (42 oC, 90s) and placed on ice for 2 min, followed by culturing into 1 ml liquid LB culture medium in 37 oC.
After 1 h, 500 µL of medium was taken and cultured on the plate of LB medium containing kanamycin (37 oC, 18 h). The recombinant plasmid was extracted and the fragment containing CRD2 and CRD3 was obtained using BamH1 and SA1 I restriction enzymes. Next, the fragment was run on agarose gel electrophoresis and then obtained using DNA extraction kit. It was then inserted into original pET28a plasmid after excision of vector by BamH1 and SA1 I restriction enzymes. The fragment was ligated to vector using T4 DNA ligase and transferred into susceptible bacteria. To verify the insertion of fragment into plasmid, the plasmid was extracted and digested by the two restriction enzymes and run on agarose gel electrophoresis.
Induction of protein expression
Isopropyl β-D-1-thiogalactopyranoside (IPTG) was used to induce the expression of recombinant protein. In pET28a vector, T7 promoter is responsible for the transcription of the inserted fragment. Although recombinant bacteria were cultured in liquid LB culture medium containing kanamycin in a shaker incubator (37 oC, 180 rpm). After 3 h, various concentrations of IPTG were added to the medium. In various time intervals of 2, 3 and 4 h after induction by IPTG, sampling was performed. The total protein was extracted by sonication on ice [13]. In addition, western blotting technique was used to detect the expression of recombinant protein.
Scale up of recombinant protein
The recombinant protein was produced on a large scale using one-liter Erlenmeyer flask and shaker incubator (37 oC, 180 rpm). Ni-NTA affinity chromatography technique was used to purify the protein. Bradford method was used to estimate the concentration of protein.
Statistical analysis
The data were statistically analyzed by one-way analysis of variance (ANOVA) using IBM Statistics SPSS software version 19, and statistical significance was set at p < 0.05. The results are expressed as mean ± standard deviation (SD, n = 3).
Results
Cloning of CRD2 and CRD3
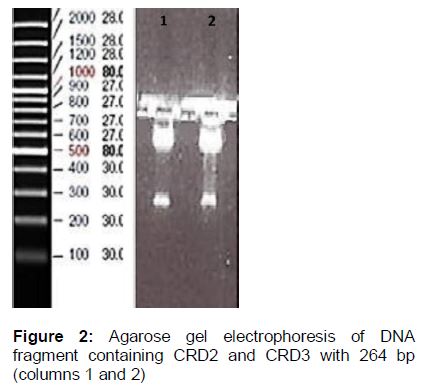
Evaluating the growth of recombinant bacteria containing CRD1, CRD2, CRD3 and CRD4 confirmed bacterial transformation (i.e. growth of antibiotic-resistance versus no growth of antibiotic-sensitive). Also, the insertion of the regions into plasmid was confirmed by agarose gel electrophoresis ().
In addition, bacterial transformation with recombinant plasmid containing CRD2 and CRD3 was confirmed. For this purpose, plasmid extraction was performed and excised by BamH1 and SA1 I restriction enzymes and then run on agarose gel electrophoresis. The fragment related to CRD2 and CRD3 was demonstrated on the gel ().
Induction of protein expression
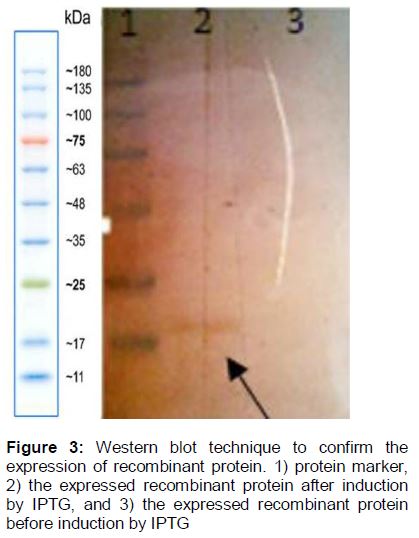
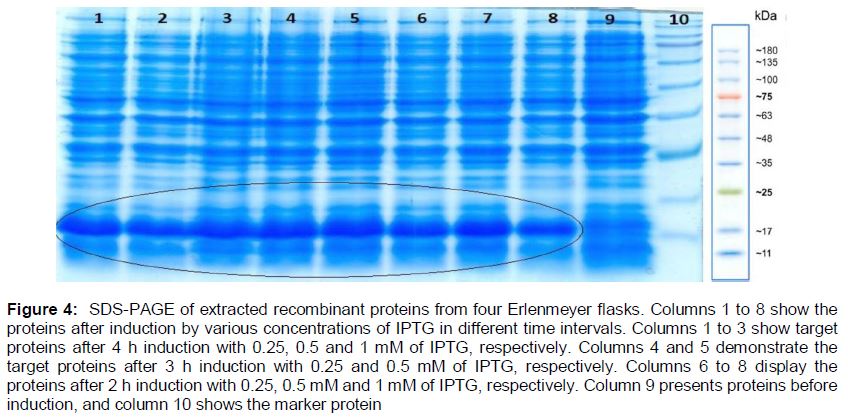
The recombinant protein was successfully expressed and detected (). In addition, the optimum condition for protein production was 3 h incubation with 0.5 mM IPTG. To extract the recombinant proteins from bacteria cultured in erlenmeyer flasks, 8 M urea and sonication (Bandelin Sonopuls HD 2070, Bandelin Elec., Germany) (3 min, 50 W) were used and the samples were run on SDS-PAGE ().
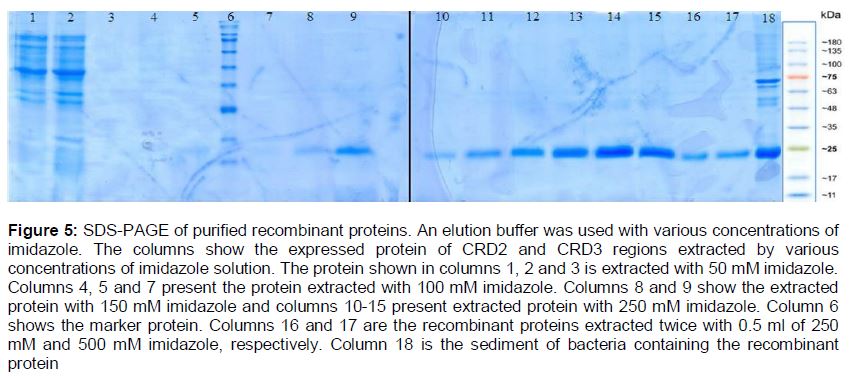
To purify the protein, Ni-NTA affinity chromatography was performed using a gradient of imidazole concentration. It was found that an elution buffer with concentration of 250 mM of imidazole was the most appropriate buffer for protein purification ().
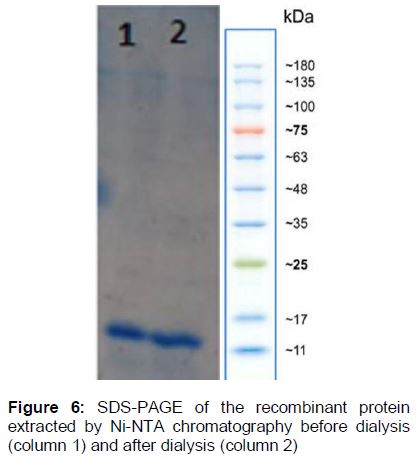
In addition, to purify the recombinant protein from urea and salt impurities, dialysis membrane (Cut off 2000 Da, Sigma) was used and the protein was run on SDS-PAGE (). Finally, evaluating the concentration of the recombinant protein by Bradford technique confirmed high yield production equal to 0.3 µg/µL.
Discussion
There are different reports on the association between TNF protein family and human disorders such as cancers, immunogenic diseases, cardiovascular, nervous system, lung and metabolic disorders [4]. TNF-α induces cell apoptosis through binding to its receptor TNFR-1 [5]. Various studies have confirmed the tumor suppressive effects of TNFR-1 [3,14]. These effects can be induced by agonistic antibodies specific for TNFR-1 [15]. Therefore, in this study we aimed to construct recombinant protein of CRD2 and CRD3 for the preparation of a specific antibody against TNFR-1. Antibodies specific for TNFR1 can be obtained through antigenic properties of receptor.
It has been demonstrated that two domains of TNFR-1, including CRD2 and CRD3, play a vital role in binding to ligands [10]. DNA coding sequence for the domains was then inserted into pET28a plasmid and transferred into competent bacteria E. coli/BL21. E. coli was chosen because of its appropriate properties for recombinant protein production such as rapid growth in simple and cheap media, well-known genetic information, availability of various types of vectors including cloning and expressing vectors, easy control and possibility of high recombinant protein production [16]. Evaluating the growth of bacteria in culture medium confirmed bacteria transformation. Also, the construction of plasmid containing CRD2 and CRD3 regions was shown by plasmid extraction and excision with BamH1 and SaI 1 enzymes and their appearance on the agarose gel electrophoresis.
In addition, the construction of the plasmid, containing CDR2 and CDR3 was reported by Jian et al [17]. They ligated the fragments to pGEX-3 X plasmid, transferred it into E. coli DH5α, and expressed it under induction of 0.25 mM IPTG [17]. Furthermore, scale up of recombinant protein production was accomplished with high yield equal to 0.3 µg/µL.
Conclusion
The recombinant protein of CRD2 and CRD3 regions of TNFR1 which can be used for the production of their specific antibodies was successfully produced. It has been observed to stimulate the expressed TNFR1 in cancer cells as well as triggers apoptotic pathways. In addition, it has been found the pET28a plasmid and E. coli/BL21 are appropriate for the production of recombinant protein, containing CRD2 and CRD3 regions of TNFR1.
Declarations
Acknowledgement
References
Archives


News Updates