Rasha Hassan Abdel-Ghany1,
Waleed Mohammed Barakat1,2,
Abdelaaty Abdelaziz Shahat3,4,
Walid El-Sayed Abd-Allah3,
Elzahraa Atef Ali1 ![]()
For correspondence:- Elzahraa Ali Email: elzahraa_atef@yahoo.com
Received: 15 February 2016 Accepted: 2 October 2016 Published: 29 November 2016
Citation: Abdel-Ghany RH, Barakat WM, Shahat AA, Abd-Allah WE, Ali EA. In vitro and in vivo hepatoprotective activity of extracts of aerial parts of Bidens pilosa L (Asteraceae). Trop J Pharm Res 2016; 15(11):2371-2381 doi: 10.4314/tjpr.v15i11.11
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the protective effects of aqueous and methanol extracts of Bidens pilosa using various in vivo and in vitro models of hepatic injury.
Methods: One kilogram of the aerial parts of Bidens pilosa was used to prepare 80 % methanol and aqueous extracts of the plant (500 g for each extract). The total phenolic content (TPC), total flavonoid content (TFC), and antioxidant activity of both extracts were evaluated. The hepatoprotective activity of these extracts in carbon tetrachloride (CCl4, 0.1 %) and D-galactosamine (700 mg/kg)-induced liver injury, respectively, was investigated in mice. Paracetamol-induced liver injury was used as in vitro reference standard.
Results: TPC and TFC of methanol extract were higher than those of the aqueous extract. The combination of methanol extract and silymarin showed the highest antioxidant activity. In vivo administration of CCl4 and D-galactosamine significantly increased the levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), and alkaline phosphatase (ALP), but decreased the total protein, albumin and glutathione (GSH) contents of liver. Co-administration of the extracts (50 mg/kg) and silymarin (100 mg/kg) effectively countered the effects of CCl4 and D-galactosamine, while also exerting their antioxidant properties. Both methanol and aqueous extracts showed hepatoprotective activity in paracetamol-induced cytotoxicity in primary cultures of rat hepatocytes.
Conclusion: Bidens pilosa possesses significant in vivo and in vitro hepatoprotective activity in mice and may be therapeutically useful as a protective agent in acute liver injury.
Introduction
The liver is a vital organ that functions as a center of metabolism and detoxification. The liver is exposed to potentially damaging substances such as microorganisms, bacterial products, or drugs delivered via the portal blood supply. Liver disease is a broad term describing a number of diseases affecting the liver, such as hepatitis, cirrhosis, cancer, and non-alcoholic fatty liver disease [1].
Liver regeneration is an example of tissue recovery after injury. Liver injury can be induced by partial hepatectomy or by various hepatotoxic chemical agents (e.g. CCl4, D-galactosamine and thioacetamide) under experimental conditions [2]. D-galactosamine has been shown to induce liver injury in rats that closely resembles the injury caused in humans by viral hepatitis, with regards to its morphological and functional features. Free radicals induced because of this injury play a major role in increasing malondialdehyde (MDA), which is an indicator of lipid peroxidation. D-galactosamine also causes alterations in lipid profile and an increase in the serum AST and ALT activities, indicating severe liver damage. Exposure to D-galactosamine has been shown to induce liver necrosis and apoptosis in rats. CCl4 is a hepatotoxic agent that induces liver damage [3]. Paracetamol is a widely used analgesic and antipyretic drug that causes acute liver damage at high doses. A growing interest has emerged globally in the identification of medicinal plants as useful therapeutic agents for the prevention of liver injury. Many synthetic drugs developed by the pharmaceutical industry have been associated with toxicity and other adverse effects. Bidens pilosa L. (family Asteraceae) is a small, erect, annual herb [4]. It is used in China as a folk medicine and as a popular ingredient in herbal teas. It has been used in the treatment of hepatitis, stomach disorders, diabetes, malaria, inflammation, and digestive disorders. Phytochemical studies of Bidens pilosa have shown the presence of alkaloids, saponins, flavonoids, polyacetylenes, and diterpenes [5]. The present study aimed to investigate the potential beneficial effects of Bidens pilosa extracts against hepatic dysfunction induced by CCl4 and D-galactosamine in mice, and by paracetamol in rat hepatic cells in vitro. The standard hepatoprotective drug silymarin was used as a positive control.
Methods
Chemicals
CCl4 was purchased from British Drug House Chemicals BDH (Poole, England). N-acetyl D-galactosamine (GaIN) was purchased from Apollo Scientific (UK). Silymarin was purchased from Chemical Industries Development (CID, Egypt). Kits for determining the AST, ALT, total proteins, and albumin were purchased from Diamond Diagnostics Inc. (Germany). Kit for determining ALP was purchased from ELITech (France). Assay kit for tissue glutathione determination was purchased from Biodiagnostic (Egypt). Other chemicals and reagents used in this study were of analytical grade.
Plant material
The aerial parts of Bidens pilosa were collected from Wadi-Hof, Helwan, Egypt. in April 2011. The authenticity of the plant was kindly confirmed by Dr. Kamal Zayed, Professor of Taxonomy, Faculty of Science, Cairo University, Egypt. A voucher specimen (no. Sh-Bp-2011) has been kept in the herbarium of Phytochemistry Department.
Preparation of Bidens pilosa methanol extract
The crude extract of the air-dried aerial parts (500 g) of Bidens pilosa was obtained by extraction with 80 % aqueous methanol three times. Extraction was continued until the extraction solvents turned colorless. The crude extracts were filtered and concentrated in vacuo at 50 °C.
Preparation of Bidens pilosa aqueous extract
Air-dried aerial parts (500 g) of Bidens pilosa were powdered and extracted with boiling distilled water for 30 min using an ultrasonic bath to ensure complete extraction. Subsequently, the extract was filtered through Whatman® filter paper No.1. The filtrates were frozen and lyophilized (LabcoNco SHEll FREEZE Dry System, England) to remove water.
Determination of total phenolic content (TPC)
80 % methanol and aqueous extracts were freshly prepared from 5 g of dried powdered aerial parts of Bidens pilosa. The 80 % methanol and aqueous extracts were prepared by diluting to 100 mL with methanol. The TPC was measured as chlorogenic acid equivalents using a UV-vis spectrophotometer. Folin-Ciocalteu method was used to determine the TPC of extracts [6]. Briefly, 1 mL of each extract was mixed with 0.5 mL of 0.2 N Folin-Ciocalteu reagent for 5 min, and 1 mL of saturated solution of Na2CO3 (40 g/100 mL) was added. After incubation at room temperature for 30 min, the reaction mixtures were diluted to 10 mL with deionized water. Absorbance of the clear solutions was measured at 725 nm. The TPC was determined from a standard calibration curve using chlorogenic acid as a standard. TPC of the extracts, expressed as mg of chlorogenic acid equivalent/g dry extract, was calculated using Eq (1).
X = 0.160Y – 0.0135 …………………….. (1)
where X = TPC concentration and Y = absorbance
Determination of total flavonoid content (TFC)
The TFC of extracts was determined using the method reported by Meda et al [6]. Briefly, 3 mL of 2 % AlCl3 in methanol and 2 mL methanol were mixed with 3 mL of 80 % methanol extract or aqueous extract for 30 min. Subsequently, the sample absorbance was measured spectrophotometrically at 415 nm against a blank sample consisting of 3 mL AlCl3 mixed with 3 mL methanol.
TFC was determined using a calibration curve, using rutin as a standard. TFC of 80 % methanol extracts, expressed as mg of rutin equivalents/g dry extract, was calculated as in Eq 2.
X = 0.151 Y + 0.00854 ……….…………… (2)
where X = TFC concentration and Y = absorbance.
Determination of free radical scavenging activity
The antioxidant activity of the extracts was determined by the 1,1-diphenyl-2-picryl-hydrazyl (DPPH) assay. Radical scavenging activity against DPPH stable radical was measured using the method of Duh et al [7]. Freshly prepared methanol solutions of DPPH (10-4 M; 2 mg DPPH dissolved in 100 mL methanol), test solutions of the extract (1 mg extract dissolved in 1 mL methanol), and reference compound Trolox (6-hydroxy-2,5,7,8-tetramethyl-chroman-2-carboxylic acid) were mixed, shaken vigorously, and kept for 15 min in the dark at room temperature.
The absorbance of the resulting solution (300 μL extract solution + 3 mL DPPH solution) was measured at 520 nm. The absorbance of a blank sample containing the same amount of methanol and DPPH solution was also measured (300 μL methanol + 3 mL DPPH solution). All determinations were performed in triplicate. The obtained results were relative to that of Trolox, which showed the highest antioxidant activity. DPPH radical scavenging activity (D) was determined using Eq 3.
D (%) = {(AB – AA)/AB}100 …………..………(3)
where AB = absorbance of blank sample and AA = absorbance of the test antioxidant after 15 min.
Isolation of rat hepatocytes and monolayer culture
A primary culture of rat hepatocytes was prepared from male Wistar rats (250 - 300 g), as reported by Kiso et al [8]. The rats were obtained from the animal house of the NRC (National Research Center, Cairo). All animal studies were approved by the Ethical Committee for Animal Handling at Zagazig University (approval no. P2 - 8 - 2015), and followed the guidelines of National Institutes of Health Guide for Care and Use of Laboratory Animals [9].
The mice were anesthetized by intraperitoneal injection of 3.4 ml/kg sodium thiopental solution (0.1 g/mL) at the time of surgery. A midline incision was made and the liver was exposed followed by cannulation of the portal vein with a needle fitted with a Teflon catheter. After the Teflon catheter was tied in place and the needle was removed, the inferior vena cava was cut below the renal vein to allow blood drainage. Subsequently, the liver was perfused with a Ca2+-free buffer (buffer 1), which contained HEPES (10 mmol/L), NaCl (137 mmol/L), KCl (2.68 mmol/L), Na2HPO4 (0.7 mmol/L), D (+) glucose (10 mmol/L), and EGTA (0.5 mmol/L), adjusted to pH 7.45 at 37 °C. The flow-rate was 11.6 ml/min. The thoracic portion of the superior vena cava was cannulated, and the inferior vena cava was tied-off above the renal vein. After perfusing the liver with buffer 1 for 15 min, the liver was perfused with a Ca2+-containing buffer (buffer 2), containing CaCl2 (5 mmol/L) and collagenase type IV (10 g/L) (pH 7.45 at 37 °C) in addition to the buffer 1 constituents, at a flow-rate of 7.5 ml/min, for 10 min. After perfusion with buffer 2, the liver was excised and placed in buffer 2, transferred under sterile conditions to laminar flow, and gently dispersed with two forceps into small pieces (2-3 mm). Subsequently, cell suspension was filtered through cotton gauze into centrifuge tubes, and centrifuged at 600 rpm for 5 min. The supernatant was aspirated and the loosely packed pellet of cells was gently resuspended in phosphate buffer (PB) (0.1 mol/L), which contained NaH2PO4 (0.2 mol/L) and Na2HPO4 (0.2 mol/L) adjusted to pH 7.6 at 37 °C, and then diluted two-folds with distilled water. The cell suspension was washed twice using this procedure. Cell viability was determined by the dye exclusion method using Trypan blue. Briefly, cell suspension (0.1 mL) was incubated with Trypan blue (0.9 mL), and the number of cells that excluded the dye and the number of cells whose nuclei were stained blue were counted using hemocytometer under a light microscope (Nikon). Complete culture medium was added to the cell suspension to reach a concentration of 4 x 106 cells/mL.
The culture medium used was RPMI-1640 medium, supplemented with 10 % heat-inactivated (56 °C for 30 min) fetal calf serum (FCS) (0.05 g/ml), 1 % penicillin-streptomycin (PS) (0.01/ml), insulin (0.7 g/mL), and dexamethasone (4 g/mL). Cells (4 x 106 cells/ml) were seeded in 96-well plates (200 µl/well) and pre-incubated in 5 % CO2-incubator at 37 °C for 22-24 h. All buffers were freshly prepared and sterilized (121 °C for 30 min) before use.
Half-maximal inhibitory concentration (IC50) determination
After pre-incubation of primary culture of rat hepatocytes for 22-24 h, the cell monolayer was checked under an inverted microscope (Olympus, CK2-TR-Japan) to confirm attachment. Cells were washed twice with phosphate buffered saline (PBS). Different concentrations (125-1000 µg/mL) of the extracts and silymarin were prepared for determination of IC50. After 2 h incubation with the extracts or silymarin, cell viability was determined by 3-(4,5- dimethyl-thiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay, as reported by Carmichael et al [10]. Absorbance of the formazan crystals produced by viable cells was measured at 580 nm using the Automatic Kinetic Microplate Reader (Labsystems Multiskan RC reader, Model 351, Finland). Each experiment was repeated three times, and the mean absorption of each concentration was calculated.
Evaluation of hepatoprotective activity
The primary rat hepatocyte monolayers were prepared and seeded as described above. Serial dilutions of the 80 % methanol extract (between 12.5-100 µg/mL) were prepared in dimethylsulphoxide (DMSO) (maximum concentration: 1 %). Cells were treated with different concentrations of the test agents. For each concentration, three replicates were used. Silymarin (50 µg/mL) was used as a positive control. The plates were incubated for 2 h, and then washed twice with PBS. Subsequently, 200 µL of 25 mM paracetamol was added to each well. After 1 h incubation with paracetamol, cell viability was determined using the MTT assay. The therapeutic index (index of the biological activity), which is the ratio of the concentration that induced toxicity and concentration that achieved hepatoprotective activity, was determined for each extract. Therapeutic index was determined as the ratio of lC50 to hepatoprotection concentration, expressed as a percentage.
Animals
The in vivo study was carried out on male albino mice (22 - 25 g) purchased from the animal house of the NRC (National Research Centre, Cairo). The mice were housed in plastic cages at room temperature, under a 12 h light – dark cycle, and allowed free access to a standard diet and tap water. All animal studies were approved by the Ethics Committee for Animal Handling at Zagazig University (ECAHZU) (No: P2 - 8 - 2015).
Determination of carbon tetrachloride (CCl4)-induced hepatotoxicity
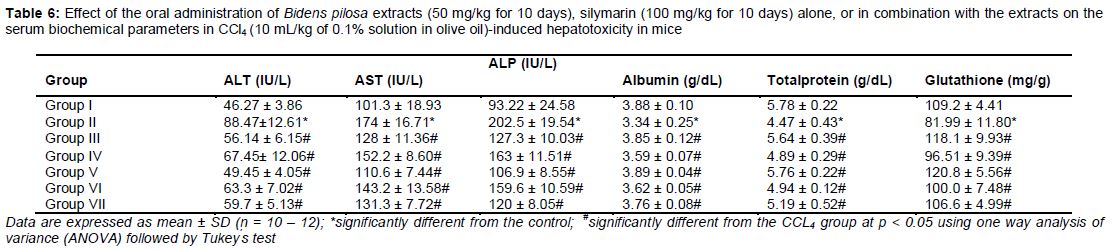
Mice were randomly divided into the following 7 groups (n = 10-12 mice per group): the control, CCl4 (10 mL/kg of 0.1 % CCl4 solution in olive oil) [11], methanol extract (50 mg/kg), aqueous extract (50 mg/kg), silymarin (100 mg/kg) [12], combination of methanol extract with silymarin, combination of aqueous extract with silymarin, and the mice group (other than the control group) intraperitoneally administered with a solution of 0.1 % CCl4 to induce acute liver injury. The animals were sacrificed 24 h after CCl4 treatment. Blood samples were collected from the orbital sinus for evaluation of biochemical parameters (ALT, AST, ALP, total protein, and albumin) and the liver tissue was isolated for the determination of reduced glutathione.
Determination of D-galactosamine (GalN)-induced hepatotoxicity
To study the effect of Bidens pilosa extracts on GalN-induced liver injury, mice were randomly divided into the following seven groups (n = 8-10): Group I – the normal control group; Group II – toxic model control group; Groups III, IV, and V – were pretreated with the methanol extract, silymarin, and aqueous extract for 10 days, respectively; Groups VI and VII – were prophylactically treated with a combination of methanol extract (50 mg/kg) and silymarin (100 mg/kg) and aqueous extract (50 mg/kg) and silymarin (100 mg/kg) for 10 days, respectively. On the tenth day, 30 min after the last administration, mice in the Groups III–VII were given an intraperitoneal injection of GalN in normal saline (700 mg/kg b.w.) [13]. After 24 h, blood samples were collected for the biochemical analysis (ALT, AST, ALP, total protein, and albumin). The liver tissue was isolated and used for the determination of reduced glutathione.
Biochemical markers of hepatic injury
Venous blood samples were collected in clean dry test tubes from the retro-orbital sinus of mice using heparinized microcapillary tubes, according to the method of Sorg & Buckner [14]. The tubes were incubated at room temperature for 15 min to allow clotting, and centrifuged at 10,000 rpm for 10 min using Heraeus Sepatech centrifuge (Labofuge 200). The serum was collected and used immediately for the determination of ALT, AST, ALP, total proteins, and albumin levels.
Tissue sampling
After animals were sacrificed, the liver was dissected, quickly frozen in liquid nitrogen, and stored at -80 °C until used for the determination of reduced glutathione.
Preparation of liver tissue homogenates
For determination of reduced glutathione level, the liver tissues were homogenized in 5 mL cold buffer (50 mM potassium phosphate, pH 7.5, and 1 mM EDTA) per gram liver tissue using a tissue homogonizer, and centrifuged at 4000 rpm for 15 min. If not assayed on the same day, the supernatant was stored at -80 °C up to one month.
Reduced glutathione level in the liver tissue was measured by a colorimetric method described by Beutler et al [15], using kits supplied by Bio-diagnostic, Egypt. The colorimetric method for measurement of the reduced glutathione (GSH) is based on the reduction of 5,5-dithio-bis-(2-nitrobenzoic acid) (DTNB) with glutathione to produce a yellow compound. The reduced chromogen is directly proportional to the GSH concentration.
Statistical analysis
All values are expressed as mean ± standard deviation (S.D). Statistical analysis was carried out using one-way analysis of variance (ANOVA) followed by Tukeyʹs post hoc multiple comparisons test using GraphPad Prism software (version 6). Differences were considered significant at p < 0.05.
Results
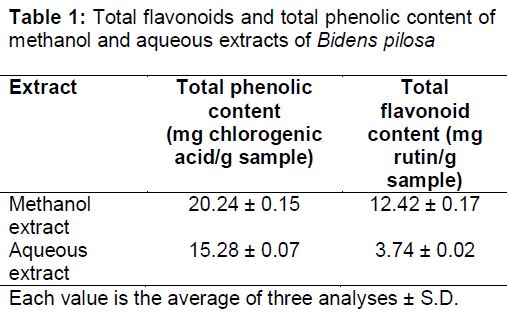
Total phenolic content (TPC)
shows that the methanol extract of Bidens pilosa contained 0.2024 mg/mL or 20.24 ± 0.15 mg chlorogenic acid per g dry extract. The aqueous extract contained 0.1528 mg/mL or 15.28 ± 0.07 mg chlorogenic acid per g dry extract.
Total flavonoid content (TFC)
As shown in , the flavonoid content of methanol extract was 0.3726 mg/3mL or 12.42 ± 0.17 mg rutin per g dry extract and of aqueous extract was 0.1122 mg/3mL or 3.74 ± 0.02 mg rutin per g dry extract.
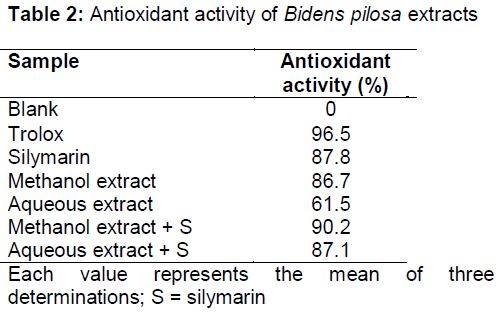
Free radical scavenging activity (DPPH)
Radical scavenging activity (expressed as absorbance percentage) of Bidens pilosa methanol and aqueous extracts are shown in (). These results showed that the antioxidant activity of methanol and aqueous extracts of Bidens pilosa (86.7 and 61.5 %, respectively) were comparable to Trolox (96.5 %) and silymarin (87.8 %). In addition, the combination of methanol or aqueous extract with silymarin showed enhanced antioxidant activity (90.2 and 87.1 % of trolox activity, respectively).
Hepatotoxicity
This assay was performed over a broad range of concentrations of the studied extracts and silymarin (from 250 - 1000 mg/mL) on monolayers of rat hepatocytes. The aqueous extracts of Bidens pilosa had IC50 at 1000 µg/mL whereas 80 % methanol extract had IC50 at a concentration >1000 µg/mL. The IC50 of silymarin was 500 µg/mL. The mean cell viabilities after exposure to 80 % methanol extract and aqueous extract at 1000 µg/mL were 85 ± 1.00 and 54.67 ± 0.58, respectively, as shown in . This indicated that both extracts exhibited high margin of safety after 2 h treatment of the rat hepatocyte monolayers ().
Hepatoprotective activity
The hepatoprotection assay was performed over a broad range of concentrations of the test samples (from 12.5 - 50 µg/mL) on monolayers of rat hepatocytes. The hepatoprotective concentration of both methanol or aqueous extracts of Bidens pilosa against the toxic effect of 25 mM paracetamol on hepatocytes was 12.5 μg/mL ().
Effect of Bidens pilosa extracts on biochemical serum parameters
As shown in , after a single injection of CCl4, the serum activities of ALT, AST, and ALP were significantly increased (p < 0.05) as compared to the normal control group. This increase indicates hepatic cell damage. Furthermore, the serum albumin, total protein, and liver glutathione contents were significantly decreased (p < 0.05) compared to the normal control group. Pre-treatment with methanol or aqueous extracts of Bidens pilosa, combination of methanol or aqueous extract with silymarin, and silymarin alone for 10 days significantly reduced the activities of serum ALT, AST, and ALP compared to the CCl4 group. It also significantly increased (p < 0.05) the serum albumin, total protein, and glutathione levels compared to the CCl4 group.
shows the effects of Bidens pilosa extracts on biochemical markers in GalN-induced hepatotoxicity in mice. Single injection of GalN significantly increased the serum activities of ALT, AST, and ALP (p < 0.05) as compared to the normal control group. This increase indicates damage of the hepatic cells. Furthermore, GaIN significantly decreased (p < 0.05) the levels of serum albumin, total protein, and liver tissue glutathione compared to the normal control mice. This acute hepatotoxicity reaction was significantly suppressed in the animals pre-treated (for 10 days) with methanol or aqueous extracts of Bidens pilosa, silymarin, and combinations of the methanol or aqueous extracts of Bidens pilosa with silymarin.
Discussion
In the absence of reliable hepatoprotective drugs in modern medicine, a large number of medicinal plants with well-established traditional use have been recommended for the treatment of liver disorders. The present study demonstrated the hepatoprotective efficacy of Bidens pilosa aerial part extracts in two in vivo animal models and one in vitro model of hepatotoxicity.
The methanol extract had high phenolic and flavonoid contents than the aqueous extract. Flavonoids are polyphenolic compounds found in plants, and are used therapeutically in many natural products. Silymarin, whose most active flavonoid component is silybin, has a powerful protective effect against CCl4-induced hepatotoxicity [16].
Previous studies on the mechanism of hepatic injury indicated that CCl4 is first metabolized by cytochrome P450 in the liver endoplasmic reticulum to the highly reactive trichloromethyl (CCl3‾) radical, which reacts readily with O2 to form the peroxytrichloromethyl (CCl3OO‾) radical. These free radicals are known to interact with cellular macromolecules, especially with unsaturated fatty acids, and initiate lipid peroxidation, thereby disrupting the structure and function of cell membrane. The present study showed that CCl4 challenge caused hepatocellular damage, which was clearly indicated by the marked elevation of serum enzyme (AST, ALT, and ALP) activities and reduction in the serum total protein, albumin, and liver glutathione content. ALT, AST, and ALP are considered the most sensitive markers for the diagnosis of hepatic injury, since they are located in cytoplasm, and deficiency of these enzymes occurs rapidly after cellular damage [17].
Results of this study are in agreement with those from previous studies that showed severe liver damage (assessed by significant elevation in ALT, AST, and ALP levels) following administration of CCl4. ALT is considered more specific for hepatic disorders because it is mainly present in the cytosol of hepatocytes but in low concentration elsewhere. AST has cytosolic and mitochondrial forms and is present in the liver, heart, skeletal muscles, brain, and pancreas. Elevation of ALT and AST levels induced by CCl4 administration may be attributed to the hepatocellular damage. In addition, the oxidative stress caused by CCl4 has been shown to induce mitochondrial dysfunction and depletion of adenosine triphosphate. CCl4 reduces glutathione content by increasing lipid peroxide generation. Furthermore, the enzymes that scavenge free radicals, such as glutathione reductase, peroxidase, and catalase are decreased during the induction of cirrhosis. Results of this study showed that the treatment with CCl4 significantly decreased the total protein and albumin levels, which is in agreement with the study of Wilkinson and Mendenhall, who found that there is a reduction in albumin synthesis in cirrhosis patients [18].
The present study showed that Bidens pilosa caused significant reduction in the activities of AST, ALT, and ALP, and significant increase in the serum total protein, albumin, and liver glutathione contents. The drugs that reduce the levels of serum marker enzymes have hepatoprotective activity. Therefore, the current study suggests that Bidens pilosa has the potential to prevent CCl4-induced hepatotoxicity.
For confirmation of the hepatoprotective activity of Bidens pilosa, we investigated whether its extracts protect against GalN-induced acute liver injury. GaIN is a well-established hepatotoxin, and induces liver injury that resembles human viral hepatitis in its morphologic and functional features. Galactosamine has great liver tendency. It disrupts the synthesis of essential uridylate nucleotides that impairs the synthesis of proteins and glycoprotein. This leads to progressive damage and change in permeability of cellular membranes, and finally enzyme leakage from hepatocytes [19].
In the present study, exposure to GaIN (single dose of 700 mg/kg, i.p.) significantly increased the plasma levels of ALT, AST, and ALP compared to the levels in control mice, which is consistent with a number of previous reports [20]. However, pre-treatment with Bidens pilosa extracts for 10 days attenuated the increased levels of ALT, AST, and ALP, and the decreased levels of albumin, total protein, and glutathione. From these results, it can be suggested that the Bidens pilosa extracts exerted hepatoprotective effect probably by stabilizing the cell membrane of hepatocytes, which prevented the loss of functional integrity and cellular leakage from the cell membrane [21]. The decreased plasma levels of ALT, AST, and ALP after exposure to Bidens pilosa extract indicate a protective effect against hepatic enzyme leakage, and therefore, hepatoprotective effect against the GaIN-induced liver damage and dysfunction in mice. Thus, it can be suggested that Bidens pilosa can preserve the structural and functional integrity of the liver against the adverse effects of D-galactosamine.
Furthermore, the decrease in plasma total proteins and albumin in the GaIN group shows the effect of GaIN on the synthetic function of hepatocytes. These results are consistent with a previous study, which reported that D-galactosamine induced inhibition of protein synthesis and secretion [22]. The reduction of serum albumin could be attributed to the reduction of albumin mRNA expression. Dabeva and Shafritz [23] found that albumin mRNA, the most abundantly expressed liver specific gene, was abruptly decreased after D-galactosamine treatment.
Bidens pilosa treatment slightly enhanced the D-galactosamine-induced reduction in the plasma levels of total proteins and albumin. The elevation induced by the aqueous extract and methanol extract was less than that induced by silymarin and was nearly similar to silymarin, respectively. The methanol extract succeeded in elevating the total protein levels and albumin to a level comparable to the normal controls. This ability to increase the levels of plasma total proteins and albumin could be attributed to the suppression of liver damage induced by D-galactosamine with some improvement of liver function.
Reactive free radicals initiate cell damage through the following two major mechanisms: covalent binding to cell membrane lipids and lipid peroxidation. In addition, oxidative stress may cause reversible or irreversible alterations of sensitive proteins that may lead to increased susceptibility to proteolytic attacks.
Glutathione is found in a remarkably high concentration in the liver and has key functions in protective processes. The reduced form of GSH is readily converted to its oxidized form (GSSG) in vivo on interaction with free radicals. In this study, D-galactosamine-induced hepatotoxicity was accompanied by a marked reduction in the hepatic GSH content, which was in agreement with previous studies, suggesting suppression of the hepatic GSH by the oxidative stress induced by D-galactosamine [24].
In vitro assay of the activity of Bidens pilosa extracts showed that for both methanol and aqueous extracts, hepatoprotective effect was achieved at 12.5 μg/mL, whereas the methanol and aqueous extracts showed hepatotoxic effect at a concentration >1000 and 1000 μg/mL, respectively. Furthermore, the therapeutic indices for methanol and aqueous extracts (> 80 and 80, respectively) were better than that for silymarin (10). This indicates that the Bidens pilosa (methanol and aqueous) extracts showed a hepatoprotective potential comparable to silymarin.
The methanol and aqueous extracts showed high therapeutic indices of > 80 % and 80 % respectively, whereas, the therapeutic index of silymarin was 10 % [25], which reflects the potent hepatoprotective activity of the tested samples. The methanol extract had higher phenolic contents than the aqueous extract. Since phenolic compounds are well known natural antioxidants, high percentage of phenolic compounds in the methanol extract could have contributed to its higher hepatoprotective activity than that of the aqueous extract.
This study demonstrated that prophylactic treatment with Bidens pilosa extracts before hepatic injury could maintain the liver enzymes at nearly normal levels, showing their potential to protect the normal functional status of the liver. The hepatoprotective activity of the Bidens pilosa extracts was probably due to their constituents (phenolic acids and flavonoids). Several previous studies have reported hepatoprotective potential of phenolic acids present in plants, such as chlorogenic and ferulic acids.
Lipid peroxidation and free radicals are involved in the main mechanisms by which hepatotoxins injure hepatocytes. The hepatoprotective activity of Bidens pilosa could be attributed to its antioxidant capacity. The antioxidant activity of Bidens pilosa extracts was assessed by measuring their DPPH scavenging activity. The extracts were found to have potent DPPH radical scavenging activity comparable to Trolox, the positive control. This effect could be due to their hydrogen donating capacity. This capacity of the extracts to scavenge DPPH radical can mediate inhibition of lipid peroxidation [26].
The methanol extract of Bidens pilosa exhibited a higher radical scavenging activity than the aqueous extract. Consequently, the combination of methanol extract of Bidens pilosa and silymarin exhibited the best antioxidant activity (90 % of Trolox activity), followed by the combination of the aqueous extract and silymarin (87 % of Trolox activity).
Conclusion
The present study shows that Bidens pilosa extracts possess significant in vivo and in vitro hepatoprotective activity. This effect may be due, at least in part, to its significant antioxidant properties. Therefore, Bidens pilosa extracts can be potentially developed into an effective protective agent against acute liver injury.
Declarations
Acknowledgement
References
Archives


News Updates